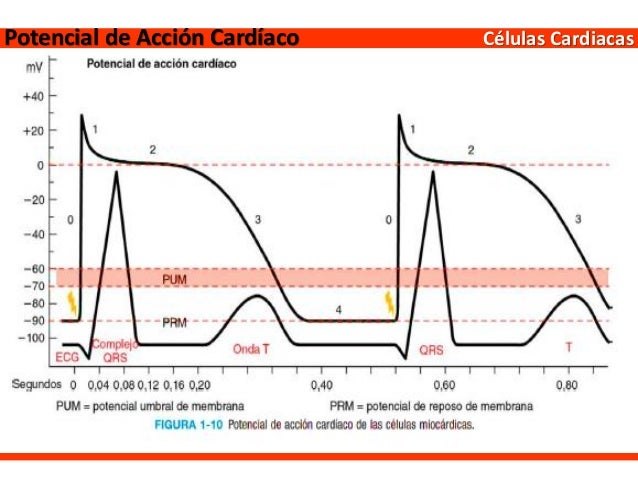
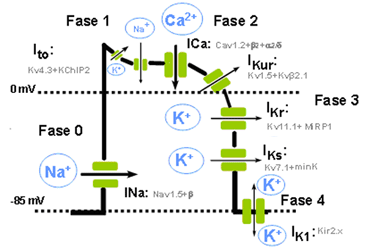
CAPÍTULO 1. Organización funcional del cuerpo humano y control del «medio interno»
CAPÍTULO 2 La célula y sus funciones
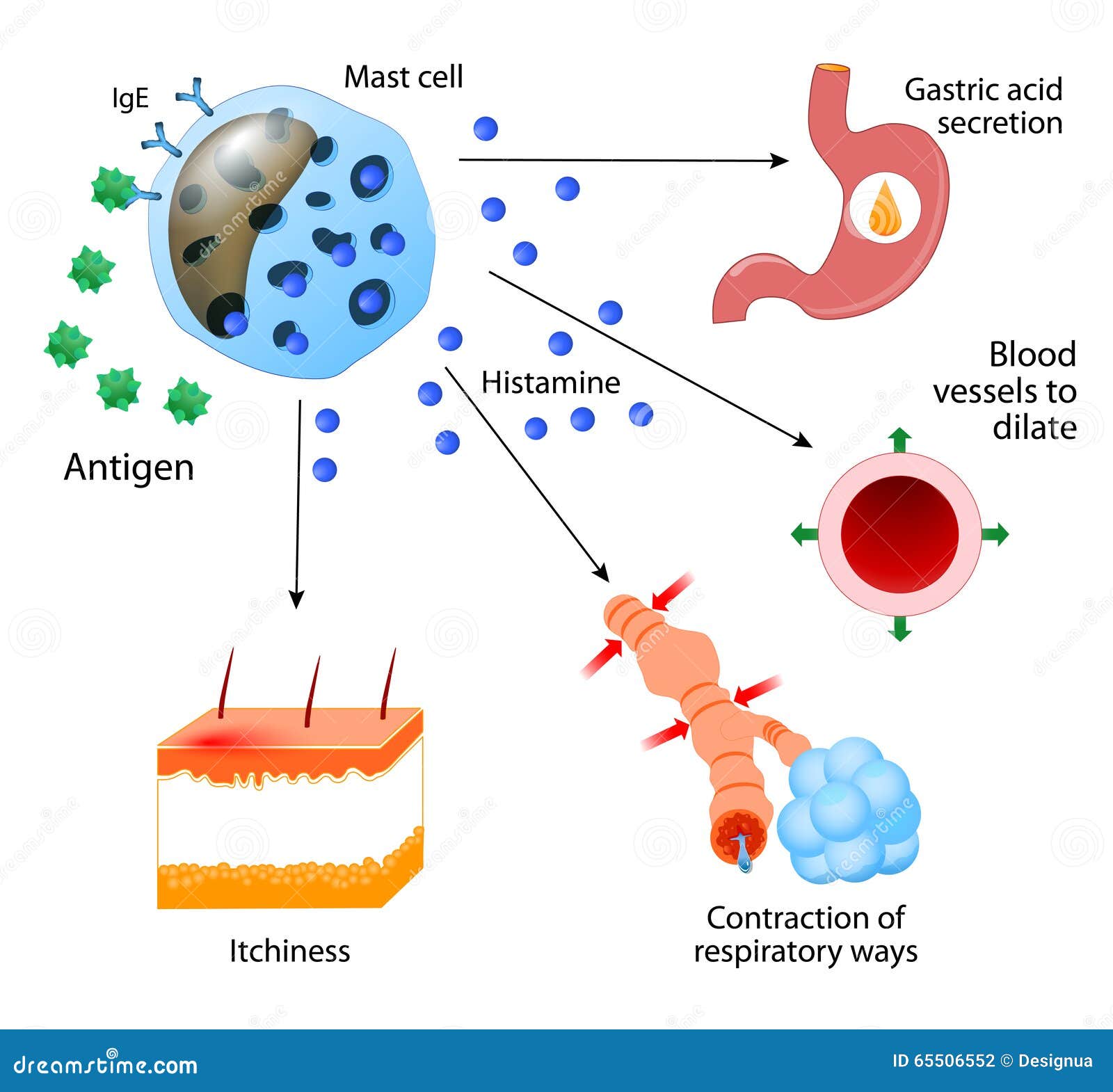
Las células son los elementos básicos que conforman el organismo; aportan la estructura de los tejidos y los órganos del cuerpo, ingieren los nutrientes y los convierten en energía, y realizan funciones especializadas.
El órgano es un conjunto de muchas células diferentes que se mantienen unidas mediante estructuras de soporte o conexiones intercelulares. Las células están especialmente adaptadas para realizar una o más funciones concretas.
Un ejemplo,
“Los eritrocitos” cuya cantidad asciende aproximadamente a 25 billones en cada ser humano, transportan el oxígeno desde los pulmones a los tejidos.
El cuerpo en su conjunto contiene en torno a 100
billones de células!
Aunque las múltiples células del cuerpo son muy diferentes entre sí, todas ellas tienen determinadas características básicas que son mayormente similares.
Organización de la célula
Sus dos partes más importantes son el núcleo y el citoplasma, que están separados entre sí por una membrana nuclear, mientras que el citoplasma está separado de los líquidos circundantes por una membrana celular que también se conoce como membrana plasmática.
Componentes de la célula
Las diferentes sustancias que componen la célula se conocen colectivamente como protoplasma.
El protoplasma está compuesto principalmente por cinco sustancias: agua, electrólitos, proteínas, lípidos e hidratos de carbono
Algunos de los iones importantes de la célula son el potasio, el magnesio, el fosfato, el sulfato, el bicarbonato y cantidades más pequeñas de sodio, cloruro y calcio
Los iones son los productos químicos inorgánicos de las reacciones celulares y además son necesarios para el funcionamiento de algunos de los mecanismos de control celulares.
Por ejemplo
los iones que actúan en la membrana celular son necesarios para la transmisión de los impulsos electroquímicos en el músculo y las fibras nerviosas.
Proteínas
las sustancias más abundantes en la mayoría de las
células son las proteínas, que normalmente constituyen entre el 10 y el 20% de
la masa celular. Son de dos tipos, proteínas estructurales y proteínas
funcionales
Las proteínas estructurales están presentes en la célula principalmente en forma de filamentos largos, los cuales son polímeros de muchas moléculas proteicas individuales. Un uso importante de este tipo de filamentos intracelulares es la formación de microtúbulos que proporcionan los «citoesqueletos» de orgánulos celulares algunos como los cilios, axones nerviosos, husos mitóticos de las células en mitosis
Las proteínas funcionales son un compuesto
habitualmente por combinaciones de pocas moléculas en un formato tubular-globular.
Estas proteínas son principalmente las enzimas de la célula, a menudo son móviles
dentro del líquido celular
Los lípidos son varios tipos de sustancias que se agrupan porque tienen una propiedad común de ser solubles en disolventes grasos. Lípidos especialmente importantes son los fosfolípidos y el colesterol, que juntos suponen solo el 2% de la masa total de la célula. Su importancia radica en que, al ser principalmente insolubles en agua, se usan para formar las barreras de la membrana celular y de la membrana intracelular que separan los distintos compartimientos celulares
En los adipocitos los triglicéridos suponen hasta el
95% de la masa celular.
Estructura física de la célula
Orgánulos
La naturaleza física de cada orgánulo es tan importante como lo son los componentes químicos para las funciones de la célula
La mayoría de los
orgánulos de la célula están cubiertos por membranas compuestas principalmente
por lípidos y proteínas
Estas membranas
son la membrana celular, la membrana nuclear, la membrana del retículo
endoplásmico y las membranas de la mitocondria, los lisosomas y el aparato de
Golgi.
Los lípidos de las membranas proporcionan una barrera
que impide el movimiento de agua y sustancias hidrosolubles desde un
compartimiento celular a otro, porque el agua no es soluble en lípidos.
Está formada casi totalmente por proteínas y lípidos,
con una composición aproximada de un 55% de proteínas, un 25% de fosfolípidos,
un 13% de colesterol, un 4% de otros lípidos y un 3% de hidratos de carbono.
El citoplasma está lleno de partículas diminutas y grandes y orgánulos dispersos. La porción de líquido gelatinoso del citoplasma en el que se dispersan las partículas se denomina citosol y contiene principalmente proteínas, electrólitos y glucosa disueltos. En el citoplasma se encuentran dispersos glóbulos de grasa neutra, gránulos de glucógeno, ribosomas, vesículas secretoras y cinco orgánulos especialmente importantes: el retículo endoplásmico, el aparato de Golgi, las mitocondrias, los lisosomas y los peroxisomas
Retículo endoplásmico
Este orgánulo ayuda a procesar las moléculas formadas por
la célula y las transporta a sus destinos específicos dentro o fuera de la célula.
Los túbulos y vesículas están conectados entre sí y sus paredes también están
formadas por membranas de una bicapa lipídica que contienen grandes cantidades
de proteínas, similares a la membrana celular
Unidas a la
superficie exterior de muchas partes del retículo endoplásmico encontramos una
gran cantidad de partículas granulares diminutas que se conocen como ribosomas.
Cuando estas partículas están presentes, el retículo se denomina retículo
endoplásmico rugoso.
Los ribosomas
están formados por una mezcla de ARN y proteínas y su función consiste en
sintetizar nuevas moléculas proteicas en la célula
Parte del retículo endoplásmico no tiene ribosomas, es lo que se conoce como retículo endoplásmico agranular, o liso. Este retículo agranular actúa en la síntesis de sustancias lipídicas y en otros procesos de las células que son promovidos por las enzimas intrarreticulares.
Aparato de Golgi
Tiene unas
membranas similares a las del retículo endoplásmico agranular y está formado
habitualmente por cuatro o más capas apiladas de vesículas cerradas, finas y
planas, que se alinean cerca de uno de los lados del núcleo.
Este aparato es prominente en las células secretoras, donde se localiza en el lado de la célula a partir del cual se extruirán las sustancias secretoras.
Lisosomas
son orgánulos
vesiculares que se forman por la rotura del aparato de Golgi y después se
dispersan por todo el citoplasma.
Los lisosomas
constituyen el aparato digestivo intracelular que permite que la célula digiera:
1) las estructuras celulares dañadas; 2) las partículas de alimento que ha
ingerido, y 3) las sustancias no deseadas, como las bacterias.
Peroxisomas
Los peroxisomas son físicamente similares a los
lisosomas, aunque difieren en dos aspectos importantes.
En primer lugar, se cree que están formados por autorreplicación (o, quizás, separándose desde el retículo endoplásmico liso) en lugar de proceder del aparato de Golgi.
En segundo lugar, contienen oxidasas en lugar de
hidrolasas. Varias de estas oxidasas son capaces de combinar el oxígeno con los
iones hidrógeno derivados de distintos productos químicos intracelulares para
formar peróxido de hidrógeno (H2O2).
El peróxido de hidrógeno es una sustancia muy oxidante que actúa junto con una catalasa, otra enzima oxidasa que se encuentra en grandes cantidades en los peroxisomas para oxidar muchas sustancias que, de lo contrario, serían venenosas para la célula.
Mitocondrias
Se conocen como los «centros neurálgicos» de la célula. Sin ellas, las células no serían capaces de extraer energía suficiente de los nutrientes y sin ellas cesarían todas las funciones celulares.
Las mitocondrias se encuentran en todas las zonas del citoplasma de la célula, pero su número total en cada célula varía de menos de cien hasta varios miles, dependiendo de la cantidad de energía que requiere la célula.
Las mitocondrias
se reproducen por sí mismas, lo que significa que una mitocondria puede formar una
segunda, una tercera, etc., siempre que la célula necesite cantidades mayores
de ATP. En realidad la mitocondria contiene un ADN similar al que se encuentra
en el núcleo de la célula.
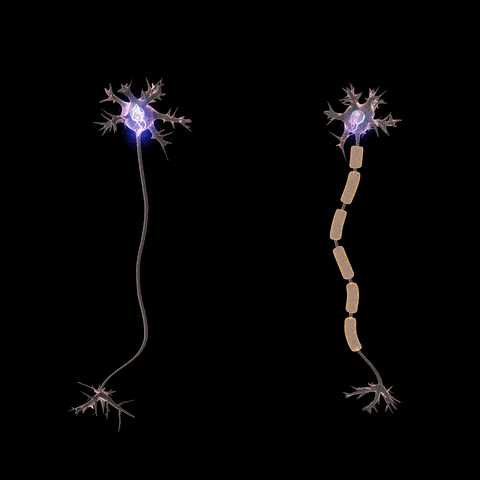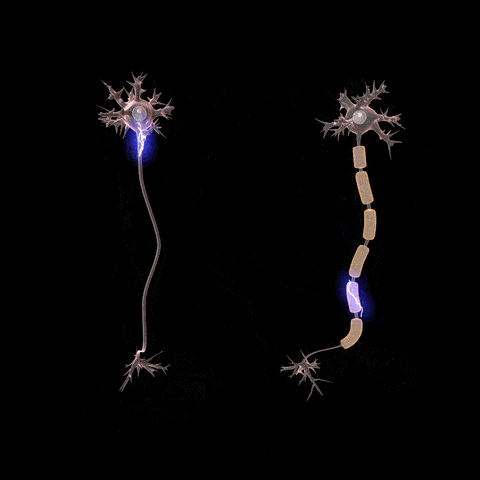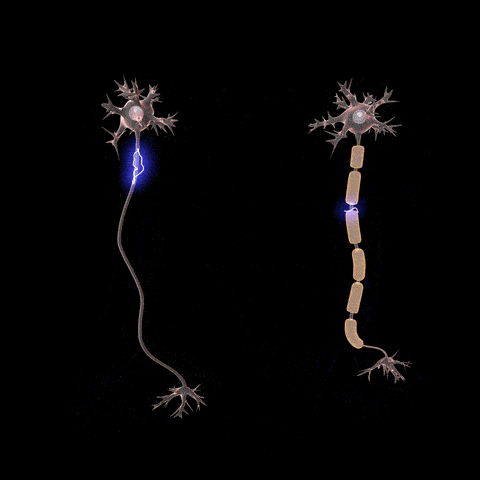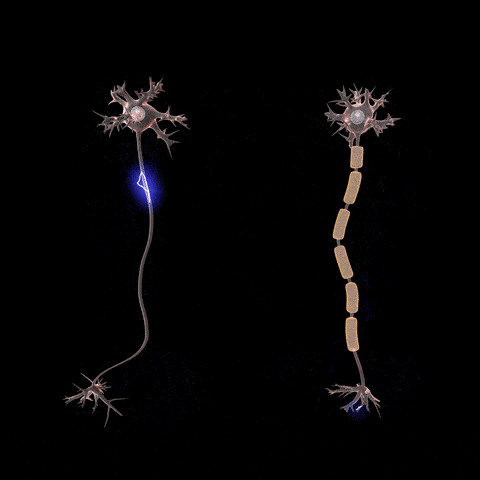
Citoesqueleto celular: estructuras filamentosas y tubulares
El citoesqueleto
celular es una red de proteínas fibrilares organizadas habitualmente en
filamentos o túbulos que se originan como moléculas proteicas precursoras sintetizadas
por los ribosomas en el citoplasma.
Es decir, una de las funciones principales de los microtúbulos es actuar como citoesqueleto, proporcionando estructuras físicas rígidas para determinadas partes de las células.
El citoesqueleto de la célula no solo determina la forma celular sino que además participa en la división de las células, permite su movimiento y proporciona una especie de ruta que dirige el movimiento de los orgánulos en el interior de las células.
Núcleo
El núcleo, que es
el centro de control de la célula, envía mensajes a esta para que crezca y
madure, se replique o muera, contiene grandes cantidades de ADN, que comprende
los genes, que son los que determinan las características de las proteínas
celulares, como las proteínas estructurales y también las enzimas
intracelulares que controlan las actividades citoplásmicas y nucleares.
Endocitocis y exocitosis
La endocitosis es una forma de transporte activo en el que una célula transporta moléculas u organismos hacia su interior, tales como proteínas.La exocitosis es todo lo contrario, la célula transporta moléculas fuera de ella.
Existen tres tipos de endocitosis: pinocitosis, fagocitosis y endocitosis mediada por receptores. La endocitosis y su contra parte, la exocitosis, son utilizadas en todas las células porque la mayoría de las sustancias químicas importantes para ellas son moléculas grandes y polares que no pueden pasar a través de la membrana, o bien la célula engulle organismos como medio de defensa y ataque.
Capítulo 25. Compartimientos del liquido corporal: líquidos extracelular e intracelular; edema.
El mantenimiento de un volumen relativamente constante y de una composición estable de los líquidos corporales es esencial para la homeostasis. Algunos de los problemas más comunes e importantes que aparecen en la medicina clínica se deben a anomalías en los sistemas de control que mantienen la constancia relativa de los líquidos corporales.
La ingestión y la pérdida de líquido están equilibradas durante las situaciones estables.
Resulta llamativa la relativa constancia de los líquidos corporales ya que hay un intercambio continuo de líquido y solutos con el ambiente externo, así como dentro de los diferentes compartimientos del cuerpo.
Ingestión diaria de agua.
El agua ingresa en el cuerpo a través de dos fuentes principales:
Se ingiere en forma de líquidos o de agua de los alimentos, que juntos suponen alrededor de 2.100 ml/día de líquidos corporales.
Se sintetiza en el cuerpo por la oxidación de los hidratos de carbono, en una cantidad de unos 200 ml/día.
Pérdida diaria de agua corporal.
Compartimientos del líquido corporal.
El líquido corporal total se distribuye sobre todo entre dos compartimientos:
Líquido extracelular (Se divide en : Líquido intersticial y plasma)
Líquido intracelular
Existe otro pequeño compartimiento de líquido que se denomina líquido transcelular:
Este compartimiento comprende el líquido de los espacios sinovial, peritoneal, pericárdico e intracelular, así como el líquido cefalorraquídeo; suele considerarse un tipo especializado de líquido extracelular, aunque en algunos casos su composición puede diferir de forma acentuada de la del plasma o de la del líquido intersticial. Todos los líquidos transcelulares constituyen alrededor de 1 a 2 l..
Compartimiento del líquido intracelular.
Unos 28 de los 42 l de líquido corporal están contenidos de los 100 billones de células y se les denomina en conjunto líquido intracelular. Por tanto, el líquido intracelular constituye alrededor del 40% del peso corporal total en una persona «media».
Compartimiento del líquido extracelular.
Todos los líquidos del exterior de las células se denominan en conjunto líquido extracelular y constituyen alrededor del 20% del peso corporal, o unos 14 l en un hombre adulto de 70 kg.
Volumen sanguíneo.
La sangre contiene líquido extracelular (el líquido del plasma) y líquido intracelular (el líquido de los eritrocitos).
El volumen sanguíneo medio de los adultos es de alrededor del 7% del peso corporal (unos 5 l). Alrededor del 60% de la sangre es plasma y el 40% son eritrocitos, pero estos porcentajes pueden variar considerablemente en diferentes personas dependiendo del sexo, el peso y otros factores.
Hematocrito (volumen del conjunto de los eritrocitos)
El hematocrito es la fracción de la sangre compuesta de eritrocitos, lo que se determina centrifugando la sangre en un «tubo de hematocrito» hasta que todas las células se acumulan en el fondo.
Dato curioso:
La composición iónica del plasma y del líquido intersticial es similar.
Debido a que el plasma y el líquido intersticial están separados solo por membranas capilares muy permeables, su composición iónica es similar.
La diferencia más importante entre estos dos compartimientos es la mayor concentración de proteínas en el plasma; debido a que los capilares tienen una permeabilidad baja a las proteínas plasmáticas, solo pequeñas cantidades de proteínas pasan a los espacios intersticiales en la mayoría de los tejidos
Medida de los volúmenes de líquido en los diferentes compartimientos hídricos del cuerpo: el principio de la dilución del indicador.
Dilución del indicador: Este método se basa en el principio de la conservación de la masa, lo que significa que la masa total de una sustancia tras la dispersión en el compartimiento líquido será la misma que la masa total inyectada en el compartimiento.
Regulación del intercambio de líquido y del equilibrio osmótico entre los líquidos intracelular y extracelular.
La distribución del líquido entre los compartimientos intracelular y extracelular, en cambio, está determinada sobre todo por el efecto osmótico de los solutos más pequeños (en especial el sodio, el cloro y otros electrólitos) que actúan a través de la membrana celular.
Principios básicos de la ósmosis y la presión osmótica
Debido a que las membranas celulares son relativamente impermeables a la mayoría de los solutos pero muy permeables al agua (es decir, son permeables selectivamente), donde quiera que haya una mayor concentración de soluto a un lado de la membrana celular, el agua se difundirá a través de la membrana hacia la región de mayor concentración de soluto. La velocidad de la difusión del agua se denomina velocidad de la ósmosis.
Osmolalidad y osmolaridad:
La concentración osmolal de una solución se denomina osmolalidad cuando la concentración se expresa en osmoles por kilogramo de agua; se llama osmolaridad cuando se expresa en osmoles por litro de solución.
Cálculo de la osmolaridad y de la presión osmótica de una solución:
Utilizando la ley de van’t Hoff, podemos calcular la posible presión osmótica de una solución suponiendo que la membrana celular es impermeable al soluto.
El equilibrio osmótico se mantiene entre los líquidos intracelular y extracelular.
Por cada miliosmol de gradiente de concentración de un soluto no difusible (uno que no atravesará la membrana celular) se ejercen unos 19,3 mmHg de presión osmótica a través de la membrana celular.
Líquidos isotónicos, hipotónicos e hipertónicos:
Líquidos isosmóticos, hiperosmóticos e hipoosmóticos:
Volumen y osmolalidad de los líquidos intracelular y extracelular en estados anormales.
Edema: exceso de líquido en los tejidos
El edema se refiere a la presencia de un exceso de líquido en los tejidos corporales. En la mayoría de los casos el edema aparece sobre todo en el compartimiento de líquido extracelular, pero puede afectar también al líquido intracelular.
Edema intracelular:
Tres procesos causan especialmente tumefacción o edema intracelular:
La hiponatremia, según se ha comentado anteriormente.
La depresión de los sistemas metabólicos de los tejidos.
la falta de una nutrición celular adecuada.
Edema extracelular:
El edema extracelular se produce cuando se acumula un exceso de líquido en los espacios extracelulares. Hay dos causas generales de edema extracelular:
La fuga anormal de líquido del plasma hacia los espacios intersticiales a través de los capilares.
La imposibilidad de los linfáticos de devolver el líquido a la sangre desde el intersticio, lo que a menudo se conoce por linfedema (incapacidad de los vasos sanguíneos de devolver líquidos y proteínas a la sangre).
Capítulo 26. El sistema Urinario: Anatomía funcional y formación de orina en los riñones.
Anatomía fisiológica de los riñones.
Organización general de los riñones y de la vía urinaria:
Los riñones se disponen en la pared posterior del abdomen, fuera de la cavidad peritoneal.
Cada riñón de un ser humano adulto normal pesa unos 150 g y tiene el tamaño aproximado de un puño cerrado.
La cara medial de cada riñón contiene una región con una muesca, llamada hilio, por la que pasan la arteria y vena renales, los linfáticos, la inervación y el uréter, que transporta la orina final desde el riñón hasta la vejiga, donde se almacena hasta que esta se vacía.
El riñón está rodeado de una cápsula fibrosa y tensa que protege sus delicadas estructuras internas.
Si se cortan los riñones de arriba abajo, las dos regiones principales que pueden verse son:
1. Corteza (externa)
2. Médula (interna)
La médula se divide en 8-10 masas de tejido en forma de cono llamadas pirámides renales.
La base de cada pirámide se origina en el borde entre la corteza y la médula y termina en la papila, que se proyecta en el espacio de la pelvis renal, una continuación en forma de abanico de la porción superior del uréter.
El borde externo de la pelvis se divide en bolsas abiertas, llamadas cálices mayores, que se extienden hacia abajo y se dividen en los cálices menores, que recogen la orina de los túbulos de cada papila.
Las paredes de los cálices, la pelvis y el uréter contienen elementos contráctiles que empujan la orina hacia la vejiga, donde se almacena hasta que se vacía en la micción.
Irrigación renal.
La nefrona es la unidad funcional del riñón.
Contiene alrededor de 800.000 a 1.000.000 de nefronas, cada una de las cuales es capaz de formar orina.
El riñón no puede regenerar nefronas nuevas.
Cada nefrona contiene:
un penacho de capilares glomerulares llamado glomérulo, por el que se filtran grandes cantidades de líquido desde la sangre.
un túbulo largo en el que el líquido filtrado se convierte en orina en su camino a la pelvis del riñón.
Micción.
La micción es el proceso mediante el cual la vejiga urinaria se vacía cuando está llena. Este proceso se realiza en dos pasos:
Primero, la vejiga se llena progresivamente hasta que la tensión en sus paredes aumenta por encima de un umbral. Esta tensión desencadena el segundo paso.
Segundo: que es un reflejo nervioso, llamado reflejo miccional, que vacía la vejiga o, si esto falla, provoca al menos un deseo de orinar. Aunque el reflejo miccional es un reflejo medular autónomo, centros presentes en la corteza cerebral o en el tronco del encéfalo pueden inhibirlo o facilitarlo.
Anatomía fisiológica de la vejiga.
Es una cámara de músculo liso compuesta de dos partes principales:
El cuerpo, que es la principal parte de la vejiga en la que se acumula la orina.
El cuello, que es una extensión en forma de abanico del cuerpo, que pasa en sentido inferior y anterior hasta el triángulo urogenital y se conecta con la uretra. La parte inferior del cuello de la vejiga también se llama uretraposterior por su relación con la uretra.
El músculo liso de la vejiga se llama músculo detrusor.
En la pared posterior de la vejiga, inmediatamente por encima del cuello de la vejiga, hay una pequeña zona triangular llamada trígono.
En la parte más inferior del vértice del trígono, el cuello de la vejiga se abre en la uretra posterior, y los dos uréteres entran en la vejiga en los ángulos más superiores del trígono.
El cuello de la vejiga (uretra posterior) tiene 2 a 3 cm de longitud, y su pared está compuesta del músculo detrusor entrelazado con una gran cantidad de tejido elástico. El músculo en esta zona se llama esfínter interno.
Más allá de la uretra posterior, la uretra atraviesa el diafragma urogenital, que contiene una capa de músculo llamada esfínter externo de la vejiga.
Capítulo 27. Filtración glomerular, flujo sanguíneo renal y su control.
Filtración glomerular: el primer paso para la formación de orina.
El primer paso en la formación de orina es la filtración de grandes cantidades de líquidos a través de los capilar es glomerular es en la cápsula de Bowman, casi 180 l al día. La mayor parte de este filtrado se reabsorbe, lo que deja únicamente 1 l aproximadamente de líquido par a su excreción al día, si bien la tasa de excreción renal de líquidos puede ser muy variable dependiendo de la ingestión de líquidos.
Composición del filtrado glomerular.
Los capilares glomerulares son relativamente impermeables a las proteínas, de manera que el líquido filtrado (llamado filtrado glomerular) carece prácticamente de proteínas y elementos celulares, incluidos los eritrocitos.
La FG es alrededor del 20% del flujo plasmático renal.
Membrana capilar glomerular.
La membrana capilar glomerular es similar a la de otros capilares, excepto en que tiene tres capas principales (en lugar de las dos habituales):
el endotelio del capilar.
una membrana basal.
una capa de células epiteliales (podocitos) rodeando a la superficie externa de la membrana basal capilar.
Determinantes de la FG.
La FG está determinada por:
La suma de las fuerzas hidrostática y coloidosmótica a través de la membrana glomerular, que da lugar a la presión de filtración neta.
El coeficiente glomerular (Kf).
Determinantes del flujo sanguíneo renal.
El flujo sanguíneo renal está determinado por el gradiente de presión a través de los vasos renales (la diferencia entre las presiones hidrostáticas en la arteria renal y en la vena renal), dividido entre la resistencia vascular total renal:
Control fisiológico de la filtración glomerular y del flujo sanguíneo renal.
Los determinantes de la FG que son más variables y están sujetos al control fisiológico son la presión hidrostática glomerular y la presión coloidosmótica capilar glomerular. Estas variables, a su vez, están influenciadas por el sistema nervioso simpático, las hormonas y los autacoides (sustancias vasoactivas que liberan los riñones y actúan a nivel local) y otros controles de retroalimentación que son intrínsecos a los riñones.
Control hormonal y por autacoides de la circulación renal.
El control
nervioso afecta principalmente a la redistribución del flujo sanguíneo hacia
las distintas zonas del organismo, el aumento de la actividad de la bomba
cardiaca y el control muy rápido de la presión arterial sistémica; este control
depende casi totalmente del sistema nervioso autónomo.
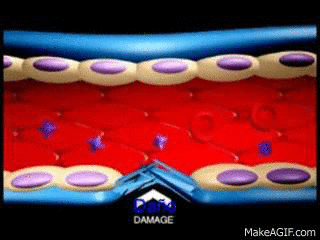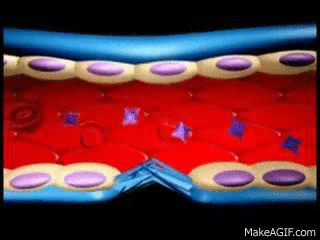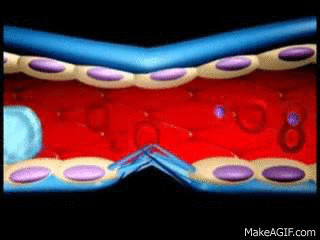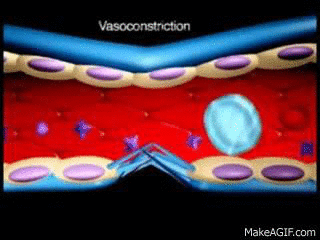
El SNA esta constituido por dos divisiones el sistema nervioso simpático; el más importante para el control de la circulación, el cual causa vasoconstricción y aumenta la frecuencia y la actividad del bombeo por el corazón; y el sistema nervioso parasimpáticoel cual ayuda en la regulación de la función cardiaca. En el sistema nervioso simpático la fibras nerviosas vasomotoras salen a través de los nervios de la columna torácica y de los dos primeros nervios lumbares, salen de la medula espinal se dirigen a las cadenas simpáticas y después por medio de dos rutas hacia la circulación:
Los nervios simpáticos que inervan la vasculatura de las viseras internas y del corazón.
2. Entrando en las porciones periféricas de los nervios raquídeos que se distribuyen hacia la vasculatura de las zonas periféricas.
La gran mayoría de los vasos sanguíneos son inervados por
estas fibras nerviosas a excepción de los capilares. La estimulación simpática
de todas las pequeñas arterias y arteriolas aumentan la resistencia vascular y
disminuye la velocidad del flujo sanguíneo a través de los tejidos, estas
fibras a su vez estimulan la actividad del corazón aumentando la frecuencia y
la fuerza de la bomba.
A diferencia del SN simpático, el SN parasimpático disminuye la frecuencia y
la actividad del bombeo del corazón, su principal función en el control de la
circulación consiste en provocar un importante descenso de la frecuencia cardiaca y también un descenso de la
contractilidad del músculo cardiaco.
Los nervios simpáticos son los encargados de transportar
una gran cantidad de fibras nerviosas vasoconstrictoras las cuales se
distribuyen hacia los segmentos de la circulación principalmente a la piel, el
intestino y el bazo.
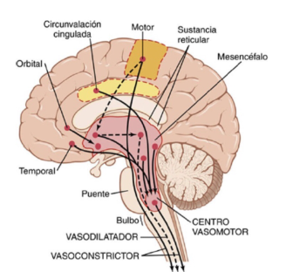
El centro vasomotores el encargado de transmitir los impulsos parasimpáticos por medio de los nervios vagos hacia el corazón, los impulsos simpáticos por medio de la medula espinal y los nervios simpáticos periféricos hacia los vasos sanguíneos del organismo.
El centro vasomotor es una zona situada
bilateralmente en la sustancia reticular del bulbo y en el tercio inferior de
la protuberancia.
Las áreas mas importantes del centro son:
Zona
vasoconstrictora: situada bilateralmente en las porciones
anterolaterales de la porción superior del bulbo, en donde las neuronas que se
originan en esta zonas secretan la noradrenalina.
Zona vasodilatadora: situada bilateralmente en las porciones
anterolaterales de la mitad inferior del bulbos, donde las fibras de estas
neuronas hacen que se inhiba la actividad vasoconstrictora de la zona C-1
provocando una vasodilatación.
Zona
sensitiva: situada bilateralmente en el núcleo del tracto solitario.
Donde las neuronas de esta zona reciben señales sensitivas por medio de los
nervios glosofaríngeos y nervios vagos pero también emiten señales
eferentes las cuales facilitan las
actividades de control de las zonas vasoconstrictoras y vasodilatadoras
consiguiéndo así el control reflejo de muchas funciones circulatorias.
Mecanismos
reflejos para mantener la presión arterial normal.
El SNA actúa para mantener la presión arterial en valores prácticamente normales a través de mecanismos reflejos de retroalimentación negativa.
Los barorreceptores es donde se inicia el
reflejo barorreceptor arterial los cuales se encuentran en las paredes de las
arterias sistémicas grandes principalmente en las paredes del seno carotideo
donde las señales se transmiten por medio de los nervios Herring hacia los nervios
glosofaríngeos y luego hacia el núcleo del tracto solitario y el cayado aórtico
que transmite señales a través de los nervios vagos hacia la misma zona del
bulbo. Los barorreceptores ya antes mencionados son encargados de controlar la
presión arterial este proceso se lleva a cabo primero con el aumento de la
presión de los vasos sanguíneos que contienen barorreceptores lo que aumenta la
descarga de los impulsos, luego las señales de estos barorreceptores entran al
núcleo del tracto solitario e inhiben el centro vasoconstrictor del bulbo y excitan el centro parasimpático
vagal. Los efectos netos son la inhibición de la actividad simpática y la estimulación de la actividad parasimpática y estos provocan vasodilatación tanto de las venas como de las arterias y el
descenso de la frecuencia cardiaca y de la fuerza cardiaca del corazón estos
factores a su vez provocan el descenso de la presión arterial como consecuencia
del descenso de la resistencia periférica y del gasto cardiaco.
Los barorreceptores ayudan constantemente a la presión arterial durante cambios postulares y las actividades
diarias, en la ausencia de los barorreceptores esenciales, las reducciones de
presión arterial disminuyen en el flujo sanguíneo cerebral provocando así la
perdida de la conciencia.
Los receptores cardiopulmonares también llamados receptores de baja presión se encuentran alojados en las paredes de la aurícula y de las arterias pulmonares los cuales son encargado de minimizar los cambios de la presión arterial en respuesta a los cambios en el volumen de la sangre, estos detectan el aumento de presión en el corazón y la circulación pulmonar, provocados por el aumento de volumen ya antes mencionado e inducen reflejos paralelos a los barorreceptores para conseguir que el sistema reflejo controle con mayor frecuencia la presión arterial. Un factor que disminuye la actividad simpática de los riñones es el estiramiento de las aurículas provocando así una dilatación refleja significativa de las arteriolas aferentes y aumentando la filtración glomerular que a su vez disminuye la reabsorción tubular del sodio, por ello el riñón excreta más sodio y agua.
Los quimiorreceptores
funcionan de una manera parecida a los barorreceptores, estos se encuentran en
dos cuerpos carotideos y en muchos cuerpos aórticos adyacentes a la aorta, estos
quimiorreceptores están formados por células quimiosensibles que excitan las
fibras nerviosas las cuales llegan a las fibras de Henring y a los nervios
vagos hacia el centro vasomotor del tronco del encéfalo. Ya que se encuentran
en contacto con la sangre arterial, siempre
que la presión arterial cae por debajo de un nivel crítico, los
quimiorreceptores se estimulan porque el descenso del flujo sanguíneo en los
cuerpos provoca la disminución del oxígeno y también la acumulación excesiva de
dióxido de carbono e iones hidrógeno, que no se eliminan debido al bajo flujo
sanguíneo. Las señales transmitidas desde los quimiorreceptores excitan el centro vasomotor, y esta eleva la presión arterial hasta la
normalidad.
Capítulo 19: Función dominante de los riñones en el control a largo plazo de la presión arterial y en la hipertensión: el sistema integrado de regulación de la presión arterial
El control de la presión arterial a corto plazo por el sistema nervioso simpático, se produce principalmente a través de los efectos del sistema nervioso sobre la resistencia vascular periférica total y la capacitancia, y sobre la capacidad de la bomba cardíaca, sin embargo, el organismo también dispone de potentes mecanismos para regular a largo plazo la presión arterial.
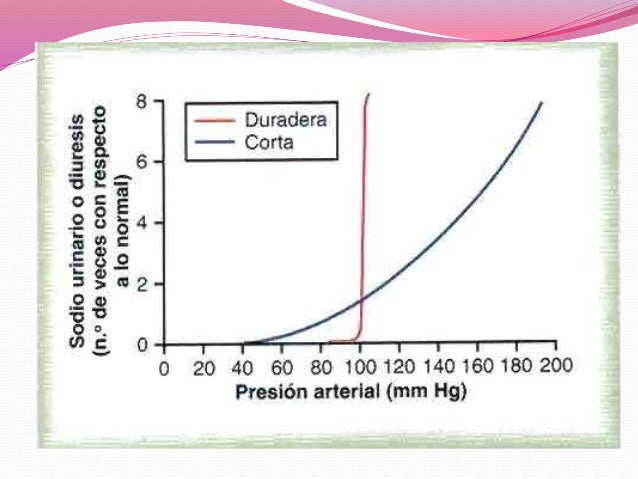
Mecanismo que se conoce como “sistema de líquidos renal-corporal”. Cuando la presión arterial aumenta en exceso, los riñones aumentan la excreción de sodio y agua debido a la nutriuresis por presión y la diuresis por presión. En consecuencia se disminuye tanto el volumen del líquido extracelular como el volumen de sangre hasta que la presión arterial vuelva a la normalidad, y los riñones excretan entonces cantidades normales de sodio y agua. Por el contrario cuando la presión arterial desciende demasiado, se reduce la excreción renal de sodio y agua. Este mecanismo puede tardar días o semanas.
La
diuresis y natriuresis se equilibran con la ingestión de sal y agua en
condiciones de equilibrio, es decir, mientras que la presión arterial este por
encima del punto de equilibrio normal, la eliminación renal es mayor que la
ingestión de sal y agua, provocando el descenso del volumen del líquido
extracelular.
Por el contrario, si la presión desciende por debajo del equilibrio normal, la eliminación renal de agua y sal es menor que la ingestión, provocando el aumento del líquido extracelular.
El
mecanismo de retroalimentación de líquidos corporales-renal muestra una ganancia por retroalimentación casi infinitaen
el control de la presión arterial a largo plazo.
Si la presión arterial desciende por debajo del punto de equilibrio, los riñones disminuyen la excreción de sal y agua hasta un nivel debajo de la ingestión, acumulándose el líquido en la sangre hasta que la presión vuelve al punto de equilibrio. Dado que el error de control de la presión es mínimo o nulo, a este sistema se le llama ganancia casi infinita.
Dos determinantes clave de la presión arterial a largo plazo son: 1) el grado de desplazamiento de la curva de eliminación renal de sal y agua y 2) el nivel de la línea de ingestión de agua y sal. No obstante si uno de ellos dos se cambia, presión arterial se regula posteriormente hasta el nuevo nivel.
El
aumento de la resistencia vascular periférica total no se puede elevar la
presión arterial a largo plazo si no se modifican la ingestión de líquidos o la
función renal. Cuando la resistencia vascular periférica total aumenta
bruscamente, presión también lo hace casi inmediatamente, pero esta no se
mantiene si la resistencia vascular de los riñones no aumenta y continua
funcionan con normalidad. Cuando la resistencia periférica y la presión
arterial aumentan, se produce diuresis por presión y la natriuresis, provocando
perdida de sal y agua. Esta pérdida continúa hasta que la presión arterial regrese
a la normalidad.
El
aumento de volumen de líquidos puede elevar la presión arterial si la capacidad
vascular no se incrementa. Esto se da en ciertos pasos, los cuales son:
El aumento del gasto cardiaco tiende por sí solo a elevar la presión arterial, si bien causa exceso de flujo sanguíneo hacia tejidos del cuerpo causando vasoconstricción, en su intento de volver a la normalidad, a lo que se llama autorregulación.
La
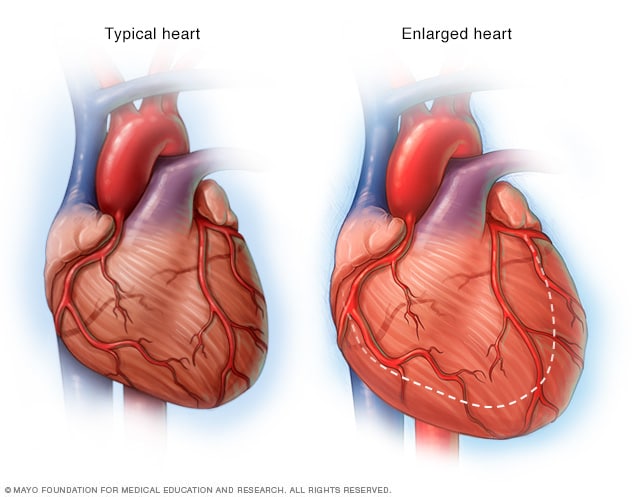
hipertensión (presión alta) se define cuando la presión diastólica es mayor de
90mmHg o la presión sistólica es mayor de 135 o 140mmHg. La hipertensión daña
la esperanza de vida, al menos de tres formas:
1) un
exceso de la carga de trabajo sobre el corazón produce insuficiencia cardiaca,
provocando un ataque cardiaco.
2) daña algún vaso sanguíneo mayor del cerebro
o hipertrofia y obstrucción final de un vaso sanguíneo cerebral.
3) siempre provoca lesiones en los riñones, produciendo insuficiencia renal.
Algunos
cambios secuenciales de la función circulatoria durante el desarrollo de la
hipertensión por sobrecarga de volumen son: debido al aumento de la ingestión
de sal y agua se produce la expansión del volumen del líquido extracelular y
volumen de sangre, el aumento de la sangre aumenta la presión media del llenado
circulatorio, retorno venoso y el gasto cardiaco, lo que provoca la elevación
de la presión arterial.
Durante
el aumento de la ingestión de sal y agua, se produce el descenso de la
resistencia periférica. Tras varios días se produce un retorno gradual del
gasto gástrico hacia la normalidad gracias a la autorregulación, lo que causa
un incremento de la resistencia periférica total. A medida que incrementa la
presión arterial, los riñones excretan el exceso mediante diuresis y
natriuesis, y se establece el equilibrio entre la ingesta y eliminación renal
de sal y agua.
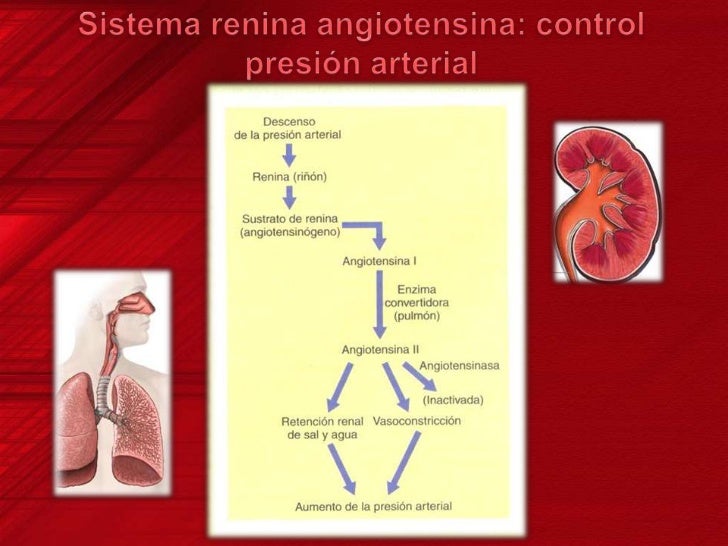
Los riñones controlan la presión a través del sistema de renina-angiotensina.
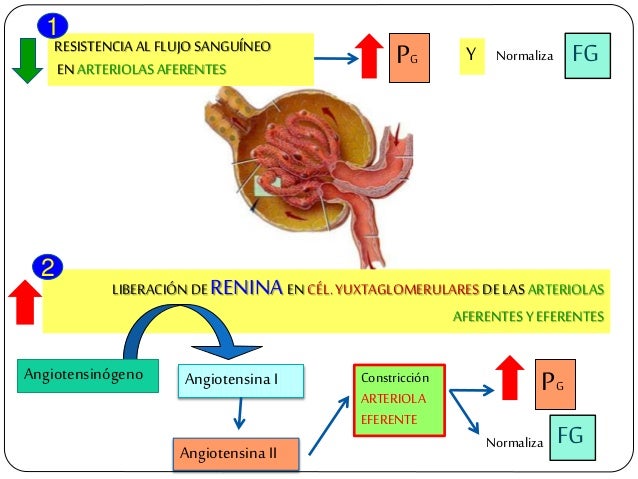
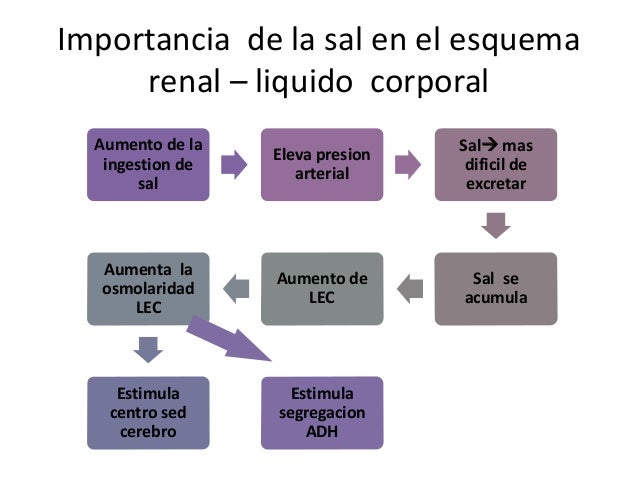
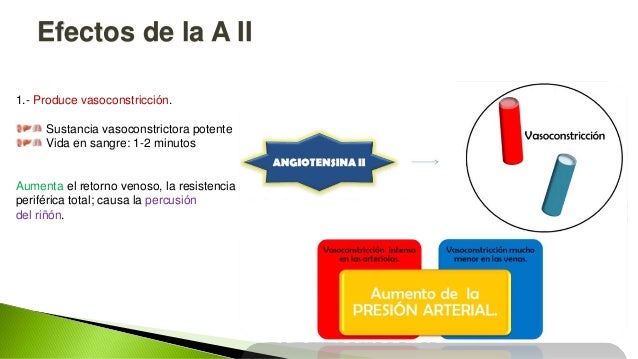
Cuando la presión arterial desciende demasiado, los riñones liberan una enzima proteica, la renina, que activa este sistema y ayuda a aumentar la presión arterial de varias formas, corrigiendo el descenso de la presión. Este sistema funciona: el descenso de la presión arterial estimula la secreción de renina y esta libera angiotensina I, la cual se convierte en II por la enzima conversora, segunda angiotensina es un vasoconstrictor y ayuda a elevar la presión, esta persiste en la sangre hasta que es inactivada por angiotensinasas. Esta segunda tiene dos funciones: contrae las arteriolas y venas de todo el cuerpo y disminuye la excreción de sal y agua.
Una de
las funciones más importantes del sistema renina-angiotensina es permitir que
la persona ingiera una pequeña cantidad sal o grande sin causar grandes cambios
en el volumen del líquido extracelular. Cuando la ingesta aumenta tanto la
presión arterial y el líquido aumentan.
Esta
mayor presión arterial también disminuye la secreción de renina y de
angiotensina II, lo que disminuye la reabsorción de agua y sal. Cuando la
ingesta de sal disminuye ocurre el efecto opuesto.
Existen
diferentes tipos de hipertensión donde interviene la angiotensina II como: la
constricción de las arterias renales causa hipertensión.
Cuando
se elimina un riñón y se coloca un elemento constrictor provoca un aumento de
presión arterial. La hipertensión producida de esta forma se conoce como
hipertensión de Goldblatt con riñón único,
el cual se debe al mecanismo de vasoconstrictor de renina-angiotensina debido
al escaso de flujo sanguíneo.
La hipertensión también puede aparecer cuando se produce la constricción de solo un riñón, mientras que la arteria del otro es normal; es a lo que se denomina hipertensión de Goldblatt con dos riñones, en este caso los dos riñones retienen sal y agua y desarrolla la hipertensión. Las causas de la hipertensión primaria (esencial) en el hombre son desconocidas. Entre el 25 y 30% de los adultos tienen presión alta. El cual la incidencia del 90% de esas personas es desconocida y este tipo es llamada hipertensión esencial. En la mayoría de los pacientes el aumento excesivo de peso y la vida sedentaria parecen desempeñar un papel importante en la causa de la hipertensión. La mayoría de los pacientes hipertensos tienen sobrepeso y en los estudios de distintas poblaciones parece demostrar que estos pueden desarrollar hasta el 65-75% del riesgo a desarrollar este problema.
Está claro que la presión arterial está regulada por varios sistemas, cada uno de los cuales realiza una función específica. Los reflejos nerviosos son mecanismos rápidos de control arterial. Los tres reflejos nerviosos que actúan con rapidez son: el mecanismo de retroalimentación de los barorreceptores, el mecanismo de isquemia en el sistema nervioso central y el mecanismo de quimiorreceptores.
Estos
no solo comienzan a reaccionar en segundos, sino que también son potentes en la
prevención del descenso agudo de la presión arterial. También actúan en la
prevención del incremento excesivo de la
presión arterial
Los
mecanismos de control de la presión arterial que actúan después de muchos
minutos son: el mecanismo de vasoconstrictor de renina-angiotensina, la
relajación de la vasculatura ante el estrés y el desplazamiento de líquidos a
través de las paredes del tejido capilar.
Los mecanismos a largo plazo para regulación de la presión arterial implican al sistema de retroalimentación de líquido renal-corporal, este mecanismo tarda varias horas en comenzar a mostrar una respuesta significativa, pero después actúa potentemente para controlar la presión arterial a lo largo de días, semanas o meses.
Uno de
los más importantes es el mecanismo de renina- angiotensina que como ya sabemos
permite que la persona tenga una ingestión de sal muy baja o muy alta con
cambios mínimos de su presión arterial.
Capítulo 20: Gasto cardíaco, retorno venoso y su regulación
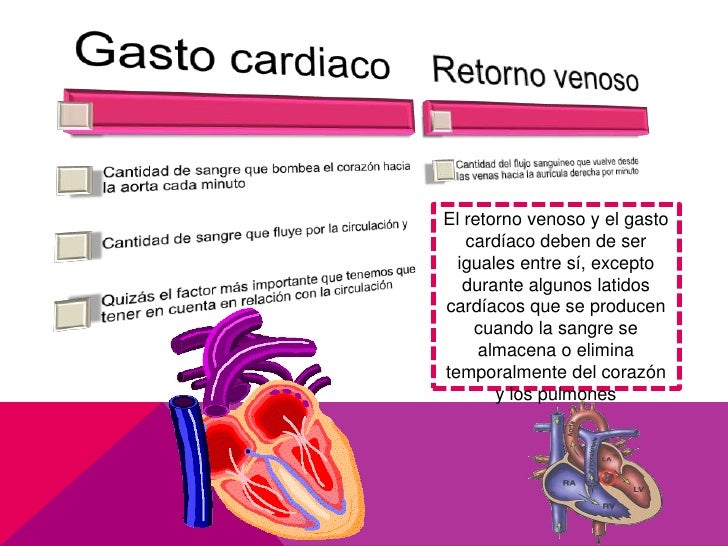
Gasto cardíaco:
Cantidad de sangre que bombea el corazón hacia la aorta cada minuto.
Cantidad de sangre que fluye por la circulación.
Suma de los flujos sanguíneos de todos los tejidos del organismo.
Retorno venoso:
Cantidad del flujo sanguíneo que vuelve desde las venas hacia la aurícula derecha por minuto.
“Deben ser iguales entre sí, excepto durante algunos latidos cardíacos que se producen cuando la sangre se almacena o elimina temporalmente del corazón y los pulmones.”
Valores
normales del gasto cardíaco en reposo y durante la actividad:
Factores
que afectan directamente al gasto cardíaco:
Nivel básico del metabolismo del organismo;
Ejercicio físico;
Edad
Tamaño del organismo.
Gasto
cardíaco en población joven y sana en reposo:
Hombres: 5.6 l/m
Mujeres: 4.9 l/m
Al aumentar la edad, disminuye la actividad corporal, por lo cual se pierde músculo esquelético, principalmente y por consecuencia, el gasto cardiaco.
Índice
cardíaco
Es el gasto
cardíaco por metro cuadrado de superficie corporal.
El gasto cardíaco aumenta en proporción a la superficie corporal.
Índice cardíaco medio normal de los adultos: 3 l/min/m2 de superficie corporal.
Efecto
de la edad en el gasto cardíaco. A los 10 años aumenta rápidamente por encima
de los 4 l/min/m2 y disminuye hasta los 2,4 l/min/m2 a los 80 años.
El
descenso del índice cardíaco indica el descenso de la actividad o de la masa
muscular con la edad.
Control
del gasto cardíaco por el retorno venoso: función del mecanismo de Frank-Starling
del corazón
El gasto cardíaco está controlado por el retorno venoso, no es el corazón propiamente quien controla el gasto cardíaco.
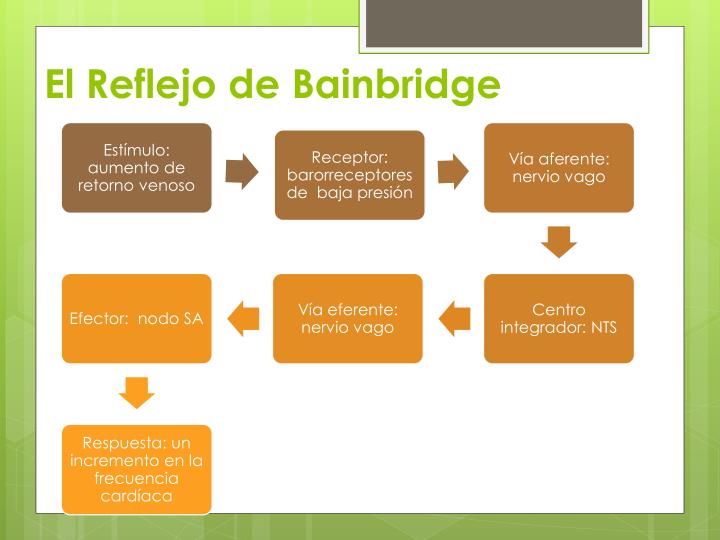
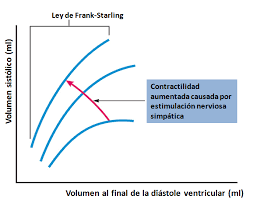
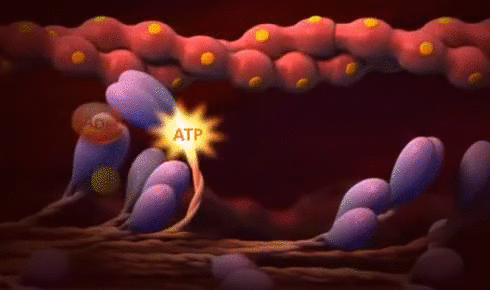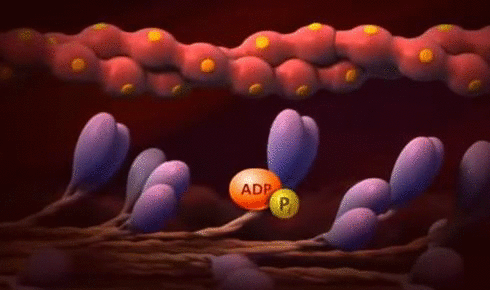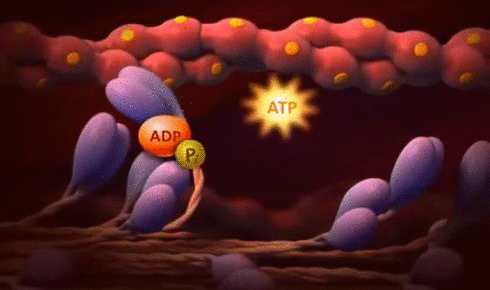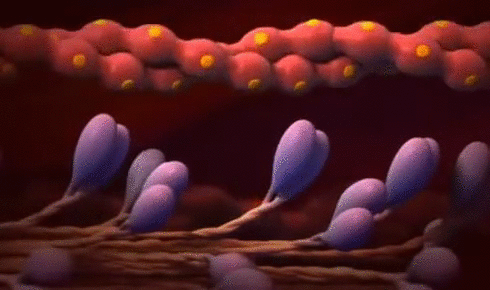
× Ley de Frank-Starling del corazón: Mecanismo propio que le permite bombear automáticamente, sin tener en cuenta la cantidad de sangre que entre en la aurícula derecha desde las venas. “Cuando aumenta la cantidad de flujo sanguíneo hacia el corazón se produce un estiramiento de las paredes de las cámaras cardíacas. Como consecuencia del estiramiento el músculo cardíaco se contrae con una fuerza mayor, por lo que vacía mejor el exceso de sangre que ha entrado desde la circulación sistémica. El estiramiento de la aurícula derecha inicia un reflejo nervioso:
® Reflejo Bainbridge:Llega primero al centro vasomotor del cerebro y después vuelve al corazón a través de los nervios simpáticos y los vagos, aumentando también la frecuencia cardíaca.
El
corazón se convierte en el factor limitante que determina el gasto cardíaco
cuando el retorno sanguíneo es mayor que el que puede bombear el corazón.
La
regulación del gasto cardíaco es la suma de laregulación
del flujo sanguíneo en todos los tejidoslocales del organismo: el
metabolismo tisularregula la mayor parte del flujo sanguíneo local
El
gasto cardíaco se encuentra determinado por la suma de todos los factores que
controlan el flujo sanguíneo local en todo el cuerpo.
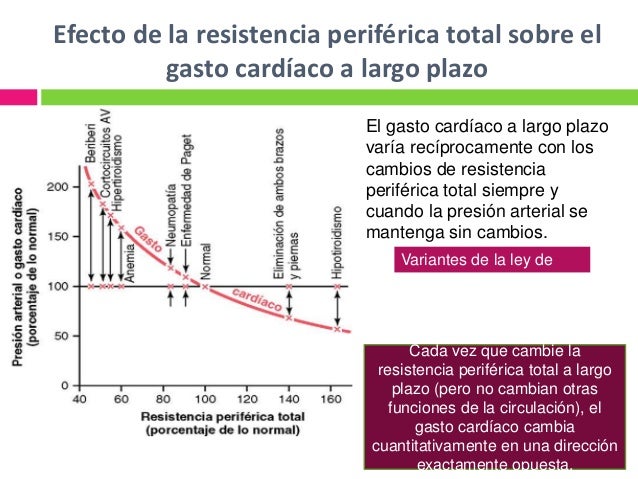
Efecto
de la resistencia periférica total sobre el gasto cardíaco a largo plazo.
El
gasto cardíaco a largo plazo varía recíprocamente con los cambios de
resistencia periférica total siempre y cuando la presión arterial se mantenga
sin cambios.
×
Cuando la resistencia periférica total es
estrictamente normal, el gasto cardíaco es normal.
×
El gasto cardíaco disminuye cuando la
resistencia periférica total aumenta por encima de lo normal;
× El gasto cardíaco aumenta cuando la resistencia periférica total disminuye.
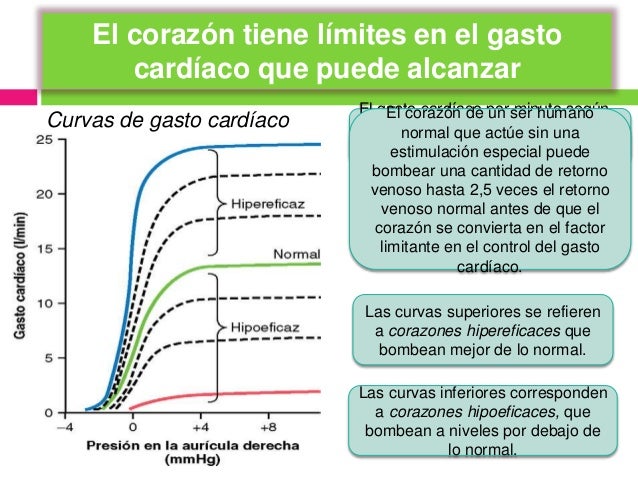
El corazón tiene límites en el gasto cardíaco que puede alcanzar
Curvas de gasto cardíaco: Miden el gasto cardíaco por minuto según cada nivel de presión en la aurícula derecha.
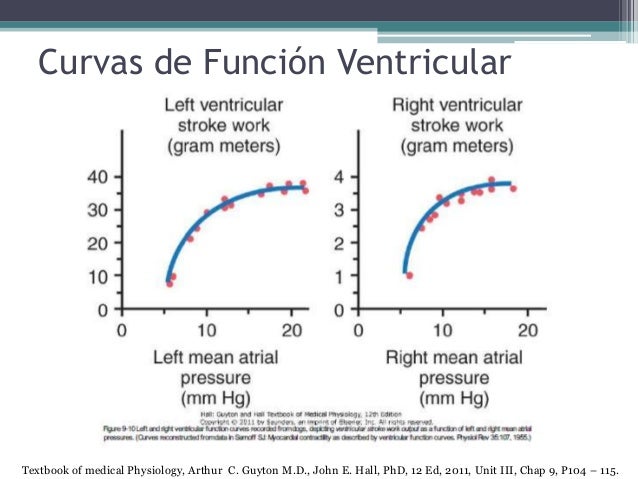
® Meseta de la curva de gasto cardíaco normal: 13 l/min, 2,5 veces el gasto cardíaco normal de 5 l/min: El corazón de un ser humano normal que actúe sin una estimulación especial puede bombear una cantidad de retorno venoso hasta 2,5 veces el retorno venoso normal antes de que el corazón se convierta en el factor limitante en el control del gasto cardíaco.
® Corazones hipereficaces: Bombean mejor de lo normal.
Factores:
1. Estimulación nerviosa: Combinación de la estimulación simpática y de la inhibición parasimpática. Aumenta el nivel de la meseta de la curva de gasto cardíaco casi hasta el doble que la meseta de la curva normal.
i.
Aumento de la frecuencia cardíaca.
ii.
Aumento de la contractibilidad del corazón.
2. Hipertrofia del músculo cardíaco: Por el aumento a largo plazo del trabajo cardíaco provoca el aumento de masa y de la fuerza contráctil, llegando a aumentar el bombeo hasta 30-40 l/min, 2.5 veces el nivel de una persona media.
® Corazones hipoeficaces: Bombean a niveles por debajo de lo normal.
×
Factores: Cualquiera que disminuya la capacidad
del corazón de bombear la sangre.
1. Aumento
de la presión arterial contra la cual debe bombear el corazón (hipertensión).
2. Inhibición
de la excitación nerviosa del corazón
3. Factores
patológicos que provocan alteraciones del ritmo cardíaco o de la frecuencia
cardíaca
4. Bloqueo
de una arteria coronaria, para provocar un «ataque cardíaco»
5. Cardiopatía
valvular
6. Cardiopatía
congénita
7. Miocarditis,
una inflamación del músculo cardíaco
8. Hipoxia cardíaca
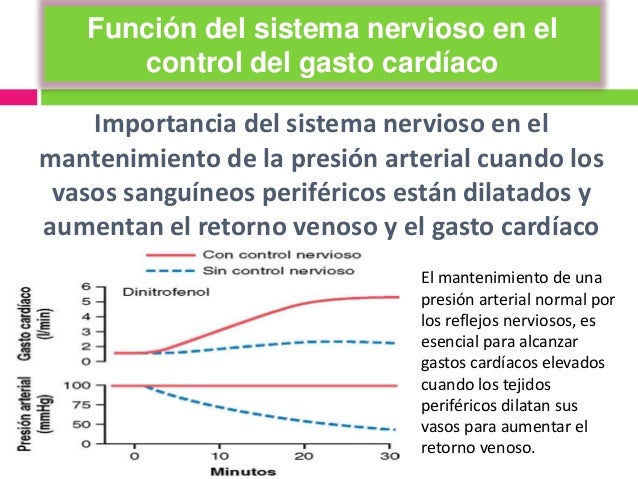
Función del sistema nervioso en el control del gasto cardíaco
En
el mantenimiento de la presión arterial cuando los vasos sanguíneos
periféricos están dilatados y aumentan el retorno venoso y el gasto
cardíaco:
® El
control nervioso previene el descenso de la presión arterial dilatando todos
los vasos sanguíneos periféricos sin causar cambios de la presión arterial pero
aumentando el gasto cardíaco casi por cuatro.
§ La vasodilatación de los vasos con dinitrofenol (líneas de puntos) provocó un descenso importante de la presión arterial hasta la mitad de lo normal, y el gasto cardíaco aumentó sólo 1,6 veces y no 4 veces.
Efecto
del sistema nervioso para aumentar la presión arterial durante el
ejercicio:
La
misma actividad cerebral que envía las señales motorasa los músculos
envía señales simultáneamente a los centrosnerviosos autónomos del
cerebro para provocar la actividadcirculatoria, provocando la
constricción de las venasgrandes y el aumento de la frecuencia y de la
contractilidad del corazón.
Elevación
y disminución patológica del gasto cardíaco
Elevación
del gasto cardíaco provocada por una reducción de la resistencia periférica
total
Son el
resultado de la reducción crónica de la resistencia periférica total.
“Cualquier factor que disminuya la resistencia periférica total crónicamente, aumentará el gasto cardíaco si la presión arterial no disminuye demasiado”.
Situaciones
que disminuyen la resistencia periférica y, al mismo tiempo, aumentan el gasto
cardíaco por encima de lo normal:
Beriberi:
×
Producida por deficiencia de tiamina (vitamina
B1) en la dieta: Disminuye la capacidad de los tejidos de usar
algunos nutrientes celulares y mecanismos del flujo sanguíneo tisular local
que, a su vez, provoquen una vasodilatación periférica compensadora.
×
La resistencia periférica total disminuye hasta
la mitad de lo normal: Los niveles de retorno venoso y gasto cardiaco a largo
plazo aumentan al doble.
Fístula arteriovenosa (cortocircuito):
× Pasa una cantidad enorme de flujo sanguíneo directamente desde la arteria hasta la vena: Aumenta el retorno venoso y el gasto cardíaco.
Hipertiroidismo:
× Metabolismo de la mayoría de los tejidos está muy aumentado: Mayor demanda de oxígeno
Liberación de vasodilatadores desde los tejidos
Reducción de la resistencia periférica total porque el flujo sanguíneo tisular reacciona por todo el cuerpo.
× Aumento del retorno venoso y gasto cardíaco del 40-80%.
Anemia:
×
Disminuye la viscosidad de la sangre por el
descenso de la concentración de eritrocitos.
×
Menor aporte de oxígeno a los tejidos, lo que
provoca vasodilatación local.
Disminución del gasto cardíaco:
Shock circulatorio: Disminución del gasto cardíaco por debajo del nivel requerido de nutrición adecuada de los tejidos.
Por anomalías que disminuyen demasiado la eficacia de la función de bomba del corazón: Nivel de bombeo por debajo de lo necesario según el flujo sanguíneo tisular que se considere adecuado.
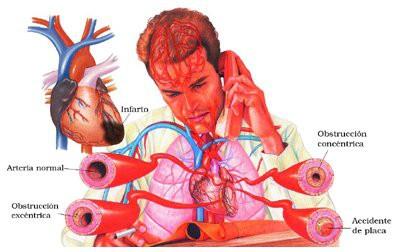
×
Bloqueo importante de los vasos sanguíneos
coronarios
×
Infarto de miocardio consecuente
×
Cardiopatía valvular grave
×
Miocarditis
×
Taponamiento cardíaco
× Alteraciones metabólicas cardíacas
× Shock cardíaco: Disminución excesiva del gasto cardíaco que causa deficiencia nutricional en todo el organismo. Muy grave, alta mortalidad.
Por factores periféricos no cardíacos: Descenso del retorno venoso:
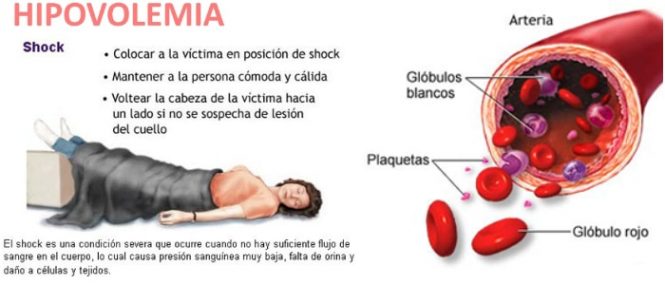
× Hipovolemia: La causa principal es hemorragia.
Disminución del llenado del corazón a niveles muy bajos ® Sangre insuficiente en los vasos sanguíneos periféricos para generar presiones vasculares suficientes para empujar la sangre de vuelta al corazón.
× Dilatación venosa aguda: Especialmente cuando el sistema nervioso simpático se vuelve súbitamente inactivo, lo cual vasodilata las venas periféricas, asentando la sangre en los vasos, evitando que regrese hacia el corazón. Ejemplo frecuente, el desmayo.
× Obstrucción de las grandes venas: Evita que la sangre vuelva al corazón.
×
Reducción de la masa tisular, en especial de
músculo esquelético:
Disminuye el consumo total de oxígeno y las necesidades de flujo sanguíneo de
los músculos.
Inactividad física
Envejecimiento normal
Reducción del ritmo metabólico de los tejidos: Dismunición del consumo de oxígeno y necesidades de nutrición de los tejidos.
Reposo en cama prolongado
Hipotiroidismo
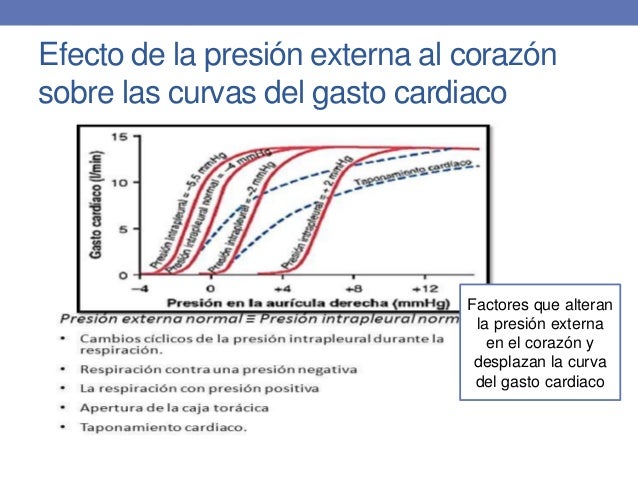
Efecto de la presión externa al corazón sobre las curvas de gasto cardíaco:
La
presión externa normal es igual a la presión intrapleural normal (la presión en
la cavidad torácica), que es de –4 mmHg.
Factores que alteran la presión externa en el corazón y desplazan la curva de gasto cardíaco:
Cambios cíclicos de la presión intrapleural durante la respiración
×
± 2 mmHg durante la respiración normal.
×
Hasta
±50 mmHg durante una respiración extenuante.
Respiración contra una presión negativa: Desplaza la presión en la aurícula derecha hacia la izquierda
Respiración con presión positiva: Desplaza la curva hacia la derecha.
Apertura de la caja torácica: Aumenta la presión intrapleural a 0 mmHg y desplaza la curva hacia la derecha 4 mmHg.
Taponamiento cardíaco: Acumulación de una gran cantidad de líquido en la cavidad pericárdica alrededor del corazón, con el aumento resultante de la presión cardíaca externa y desplazamiento de la curva hacia la derecha.
Combinaciones de los distintos patrones de curvas de gasto cardíaco
Lacurva
final del gasto cardíaco cambia como consecuencia de:
Cambios simultáneos de la presión cardíaca externa.
Eficacia del corazón como bomba.
Corazón hipereficaz + Aumento de la presión intrapleural = Incremento del nivel máximo de gasto cardíaco debido a la mayor capacidad de bombeo del corazón. La curva de gasto cardíaco aparecería desplazada hacia la derecha (hacia presiones auriculares más elevadas) a causa del aumento en la presión intrapleural.
Ruidos cardíacos y soplos
S1: – Causado por la
sístole ventricular, cierre de la válvula AV. El “tum” se relaciona con el
cierre de las válvulas auriculoventriculares (AV) al comienzo de la sístole
(que aquí es cuando el corazón se exprime y saca la sangre que tenía
contenida).
S2: – Es más breve y agudo
que el primero – Lo ocasionan el cierre valvular aórtico y el sigmoideo
pulmonar. El “ta” se asocia con el cierre de las válvulas semilunares (aortica
y pulmonar) al final de la sístole (aquí es cuando toda la sangre que tenía el
corazón terminó de salir y se vuelve a llenar para iniciar un nuevo ciclo)
S3: – Se aprecia en
diástole por disfunción ventricular. Éste es débil y retumba. Se aprecia al
comienzo del tercio medio de la diástole y se ausculta sólo en el foco
apexiano. Es habitual en la niñez, frecuente en adolescentes y muy raro después
de los 40 años.
S4: – Es un ruido auricular debido a la tensión de las válvulas AV como del miocardio ventricular, debido a un llenado acelerado. También se conoce como ruido auricular. Es ocasionado por un llenado rápido. No se ausculta en personas menores de 50 años. Puede tener origen derecho o izquierdo y sus causas son:
Derecho:
-Hipertensión pulmonar
-Cardiomiopatía
-Estenosis valvular
pulmonar
Izquierdo:
-Hipertensión sistémica
-Estenosis aórtica
valvular
-Cardiomiopatía
-Cardiopatía isquémica
Quinto ruido (cuando ya no
se escucha nada), éste último se correlaciona con la presión arterial
diastólica (aproximadamente 80 mmHg).
Soplos:
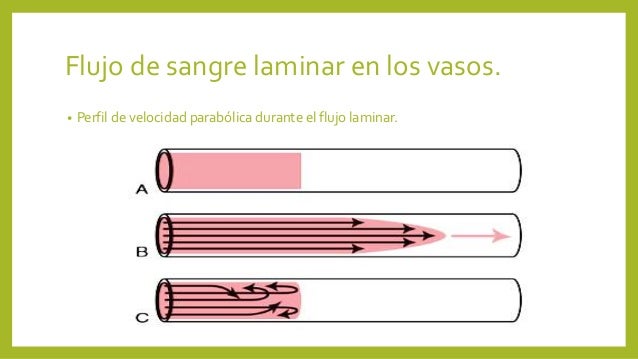
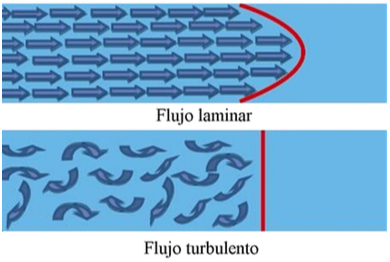
Los líquidos (en este caso
la sangre) tienen una circulación laminar unidireccional; esto se puede
entender mejor al recordar la manera en que desliza un jugador de póker las
cartas sobre la mesa, al inicio se tiene una torre de naipes, luego el jugador
desliza esta torre expandiéndola en la mesa, de tal forma que las cartas que
tocan la mesa se deslizarán de una forma más lenta, que las cartas que se
deslizan sobre ellas, la cuáles lo hacen a mayor velocidad.
La sangre, al circular
dentro de los vasos sanguíneos, presenta un comportamiento similar, la que se
encuentra en contacto con la pared del vaso casi no se desliza, sin embargo, la
sangre que corre en medio del vaso se desliza rápidamente sobre la antes
mencionada y forma un flujo laminar unidireccional. Cuando la sangre encuentra
un vaso con un diámetro que disminuye bruscamente, se interrumpe el flujo
laminar, y el flujo fluye en todas direcciones, lo que crea turbulencia y da
lugar a una especie de remolino dentro del segmento estrecho (flujo
turbulento). Este flujo turbulento produce un sonido que puede ser audible
(soplo) y que presentará diferentes tonalidades que dependerán de lo estrecho
de la luz del vaso por donde circula la sangre, éste es el fenómeno que
describió Korotkoff.
Ejemplos especiales del control «metabólico» a corto plazo del flujo sanguíneo local
Los mecanismos para controlar el flujo sanguíneo local se denominan «mecanismos metabólicos» porque todos ellos funcionan en respuesta a las necesidades metabólicas de los tejidos.
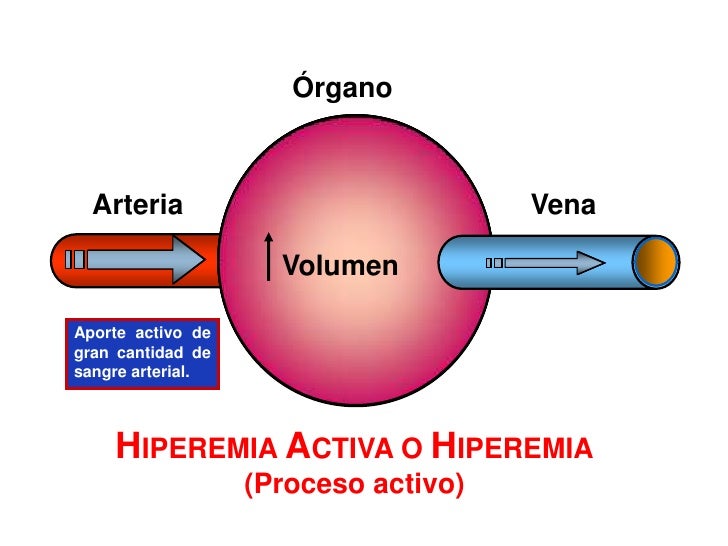
Hay otros dos ejemplos especiales de control metabólico del flujo sanguíneo local, la hiperemia reactiva y la hiperemia activa.
Hiperemia reactiva. Cuando la sangre que irriga un tejido se bloquea durante unos segundos durante 1 h o más, y después se desbloquea, el flujo sanguíneo que atraviesa el tejido aumenta inmediatamente hasta 4-7 veces con respecto a lo normal; este aumento del flujo continuará durante varios segundos, si el bloqueo ha durado sólo unos segundos, pero a veces continuará muchas horas, si el flujo sanguíneo ha estado interrumpido durante 1 h o más. Este fenómeno se conoce como hiperemia reactiva.
La hiperemia reactiva es otra manifestación del mecanismo de regulación «metabólico» del flujo sanguíneo, es decir, la falta de flujo pone en marcha todos estos factores que provocan la vasodilatación. Después de períodos cortos de oclusión vascular la cantidad extra de flujo sanguíneo que aparece durante la fase de hiperemia reactiva dura lo suficiente como para reponer casi exactamente el déficit de oxígeno tisular que se ha acumulado durante el período de oclusión. Este mecanismo resalta la estrecha conexión existente entre la regulación del flujo sanguíneo local y el aporte de oxígeno y de otros nutrientes a los tejidos.
Hiperemia activa. Cuando cualquier tejido se vuelve muy activo, como un músculo que hace ejercicio, una glándula gastrointestinal durante el período hipersecretor o incluso en el cerebro durante la actividad mental rápida, la velocidad del flujo sanguíneo aumenta a través del tejido. En este caso, de nuevo, se puede entender fácilmente esta hiperemia activa si se aplican los principios básicos de control del flujo sanguíne local. El incremento del metabolismo local hace que las células devoren rápidamente los nutrientes del líquido tisular y también que liberen grandes cantidades de sustancias vasodilatadoras. El resultado es que se dilatan los vasos sanguíneos locales y, por tanto, aumenta el flujo sanguíneo local. De esta forma, el tejido activo recibe los nutrientes adicionales necesarios para mantener este nuevo nivel de funcionamiento. Como ya hemos mencionado, la hiperemia activa del músculo esquelético aumenta el flujo sanguíneo muscular local hasta en 20 veces durante el ejercicio intenso.
«Autorregulación» del flujo sanguíneo cuando la presión arterial cambia de la normalidad: mecanismos «metabólicos» y «miógenos»
En cualquier tejido del organismo el rápido incremento de la presión arterial provoca un aumento inmediato del flujo sanguíneo, pero en menos de 1 min ese flujo volverá a la normalidad en la mayoría de los tejidos, incluso aunque la presión arterial se mantenga elevada. Esta normalización del flujo se denomina «autorregulación del flujo sanguíneo».
Una vez se haya producido esta autorregulación, el flujo sanguíneo local de la mayoría de los tejidos del organismo estará relacionado con la presión arterial, aproximadamente según la curva «aguda» de trazo continuo Obsérvese que entre una presión arterial de 70 mmHg y otra de 175 mmHg se produce un aumento del flujo sanguíneo sólo del 20 al 30%, incluso cuando la presión arterial aumente en un 150%. Durante casi un siglo se han mantenido dos opiniones que explicarían el mecanismo de autorregulación a corto plazo, la teoría metabólica y la teoría miógena.
La teoría metabólica se puede entender fácilmente si se aplican los principios básicos de regulación del flujo sanguíneo local que hemos comentado en las secciones previas. Es decir, cuando la presión arterial es demasiado elevada, el exceso de líquido proporciona demasiado oxígeno y demasiados nutrientes de otro tipo hacia los tejidos y «lava» los vasodilatadores liberados por los tejidos. Estos nutrientes (en especial, el oxígeno), junto con el descenso en los niveles tisulares de vasodilatadores, provocan entonces la constricción de los vasos sanguíneos y el retorno del flujo casi a la normalidad, a pesar de que aumente la presión. No obstante, según esta teoría miógena habría otro mecanismo no relacionado con el metabolismo tisular que explicaría el fenómeno de la autorregulación.
Esta teoría se basa en la observación de que el estiramiento brusco de los vasos sanguíneos pequeños provoca la contracción del músculo liso
Por tanto, se ha propuesto que cuando una presión arterial elevada estira el vaso se provoca, a su vez, una constricción vascular reactiva que reduce el flujo sanguíneo casi a la normalidad. Por el contrario, con presiones bajas el grado de estiramiento del vaso es menor, por lo que el músculo liso se relaja, reduce la resistencia vascular y ayuda a recuperar la normalidad del flujo.
La respuesta miógena es inherente al músculo liso vascular y puede producirse en ausencia de influencias nerviosas u hormonales. Es más pronunciada en las arteriolas, pero se puede ver también en arterias, vénulas, venas e incluso en vasos linfáticos.
La contracción miógena se inicia por la despolarización vascular inducida por el estiramiento, que tiende a aumentar rápidamente la entrada de ion calcio desde el líquido extracelular hacia las células, provocando su contracción. Los cambios de la presión vascular también pueden abrir o cerrar otros canales iónicos que influyen en la contracción vascular. Se desconocen los mecanismos por los cuales los cambios de la presión provocan la apertura o el cierre de los canales iónicos vasculares, pero es probable que consistan en efectos mecánicos de la presión sobre las proteínas extracelulares que están ancladas en los elementos del citoesqueleto de la pared vascular o en los propios canales iónicos. El mecanismo miógeno parece ser importante para prevenir el estiramiento excesivo del vaso sanguíneo cuando aumenta la presión sanguínea. No obstante, también se desconoce la importancia de este mecanismo en la regulación del flujo sanguíneo porque este mecanismo de detección de la presión no puede detectar directamente los cambios del flujo sanguíneo en el tejido. En realidad, los factores metabólicos anulan el mecanismo miógeno cuando las demandas metabólicas de los tejidos están significativamente aumentados, como durante el ejercicio muscular enérgico, lo que provoca un incremento espectacular del flujo sanguíneo en el músculo esquelético.
Mecanismos especiales del control a corto plazo del flujo sanguíneo en tejidos específicos
Aunque los mecanismos generales de control del flujo sanguíneo local que hemos comentado hasta ahora actúan en la mayoría de los tejidos del organismo, en algunos territorios especiales actúan otros mecanismos totalmente diferentes. Todos los mecanismos se comentarán en este texto en relación con cada órgano específico, pero hay dos que merecen una atención especial:
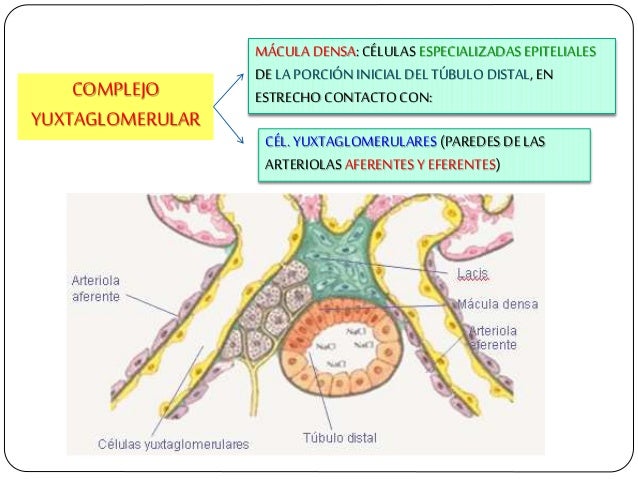
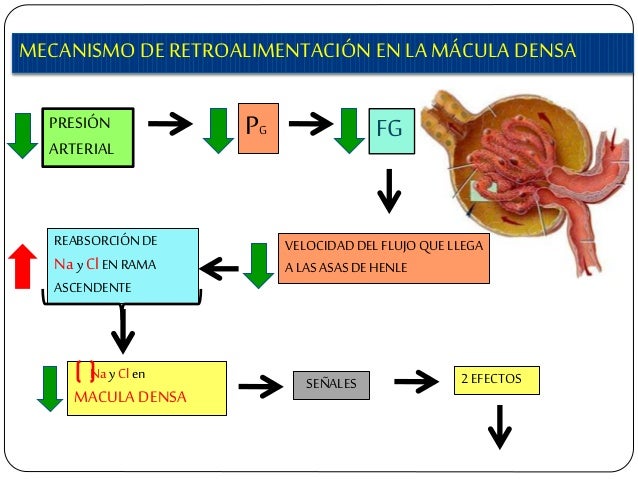
En los riñones, el control del flujo sanguíneo se basa en gran medida en un mecanismo denominado retroalimentación tubuloglomerular, en el que una estructura epitelial del túbulo distal, la mácula densa, detecta la composición del líquido al inicio de dicho túbulo. La mácula se sitúa en la zona en que el túbulo distal se encuentra cerca de las arteriolas aferente y eferente del aparato yuxtaglomerular de la nefrona. Cuando se filtra demasiado líquido de la sangre a través del glomérulo hacia el sistema tubular, las señales de retroalimentación de la mácula densa provocan constricción de las arteriolas aferentes, reduciendo de esta forma tanto el flujo sanguíneo renal como la tasa de filtración glomerular a valores normales o casi normales.
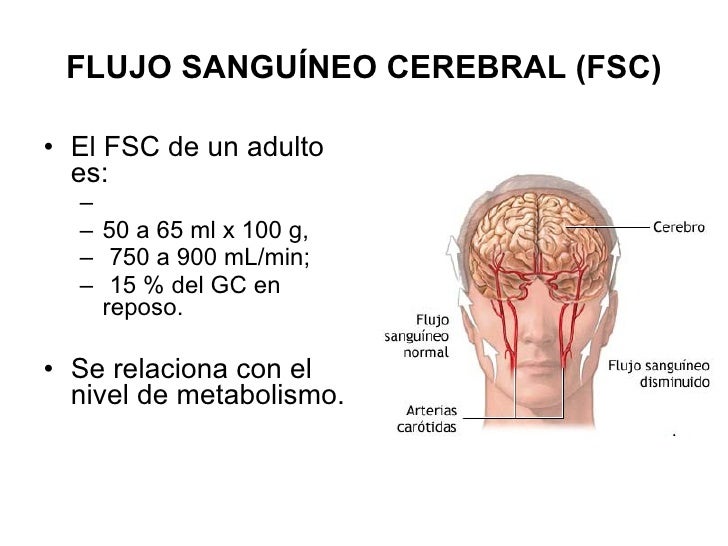
En el cerebro, además del control del flujo sanguíneo dependiente de la concentración de oxígeno tisular, las concentraciones de dióxido de carbono y de iones hidrógeno tienen una gran importancia. El aumento de cualquiera de ellos dilata los vasos cerebrales y permite el lavado rápido del exceso de dióxido de carbono o de iones hidrógeno de los tejidos cerebrales, lo que es importante porque el nivel de excitabilidad del propio cerebro depende en gran medida del control exacto de las concentraciones de dióxido de carbono y del ion hidrógeno.
En la piel, el control del flujo sanguíneo está relacionado estrechamente con la regulación de la temperatura corporal. El flujo cutáneo y subcutáneo regula la pérdida de calor del cuerpo mediante la determinación del flujo de calor desde el centro a la superficie del organismo, donde se pierde calor hacia el medio exterior. El flujo sanguíneo en la piel está controlado en gran medida por el sistema nervioso central a través de los nervios simpáticos
Aunque el flujo sanguíneo en la piel supone sólo en torno a 3 ml/min/100 g de tejido en tiempo frío, cuando se necesita pueden producirse cambios importantes con respecto a estos valores. Cuando los seres humanos se exponen a un calentamiento, el flujo sanguíneo cutáneo puede incrementarse muchas veces, hasta 7 u 8 l/min para todo el organismo. Si se reduce la temperatura corporal, el flujo sanguíneo en la piel disminuye, para descender a poco más de cero a temperaturas muy bajas. Incluso con una vasoconstricción acusada, el flujo sanguíneo cutáneo suele ser suficientemente elevado para satisfacer las demandas metabólicas básicas de la piel.
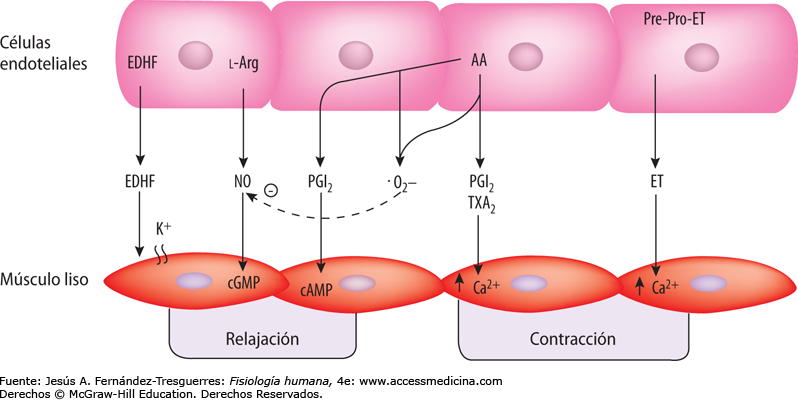
Control del flujo sanguíneo tisular por medio de factores de relajación y contracción de origen endotelial
Las células endoteliales que recubren los vasos sanguíneos sintetizan varias sustancias que, cuando se liberan, afectan al grado de relajación o contracción de la pared arterial. Para muchos de estos factores de relajación o contracción de origen endotelial, las funciones fisiológicas apenas se están empezando a comprender y, en la mayoría de los casos, todavía no se han desarrollado aplicaciones clínicas.
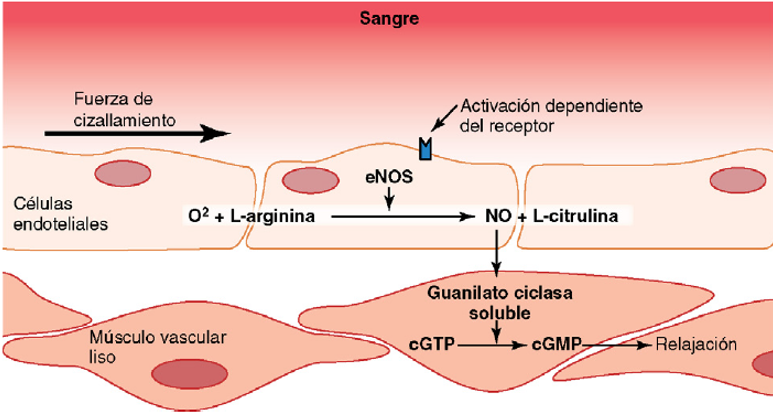
Óxido nítrico: un vasodilatador liberado por células endoteliales sanas. El más importante de los factores de relajación de origen endotelial es el óxido nítrico (NO), un gas lipófilo que es liberado por las células endoteliales como respuesta a diversos estímulos químicos y físicos. La enzima óxido nítrico sintasa (NOS) de las células endoteliales sintetiza el NO a partir de arginina y oxígeno y por reducción de nitrato inorgánico. Después de la difusión fuera de la célula endotelial, el NO tiene una semivida en sangre de sólo 6 s, aproximadamente, y actúa principalmente en los tejidos locales en los que es liberado. El NO activa las guanilato ciclasas solubles en las células de músculos lisos vascular lo que produce la conversión de trifosfato de guanosina cíclico (cGTP) a monofosfato de guanosina cíclico (cGMP) y la activación de proteína cinasa dependiente de cGMP (PKG), que tiene varias acciones que provoca la relajación de los vasos sanguíneos. Cuando la sangre circula a través de las arterias y arteriolas provoca fuerzas de cizallamiento sobre las células endoteliales por el arrastre viscoso de la sangre contra las paredes vasculares. Esta tensión contorsiona las células endoteliales en la dirección del flujo y provoca un incremento significativo de la liberación de NO. El NO relaja entonces los vasos sanguíneos, lo que es una ventaja porque los mecanismos metabólicos locales para el control del flujo sanguíneo tisular dilatan principalmente las arterias y arteriolas muy pequeñas en cada tejido. Aun así, cuando aumenta el flujo sanguíneo a través de una porción microvascular de la circulación, estimula de forma secundaria la liberación de NO de los grandes vasos debido al aumento del flujo y a la tensión de cizallamiento en estos vasos.
El NO liberado aumenta los diámetros de los grandes vasos sanguíneos proximales siempre que el flujo sanguíneo microvascular aumenta distalmente. Sin esta respuesta disminuiría significativamente la eficacia del control del flujo sanguíneo local, porque una parte significativa de la resistencia al flujo sanguíneo se produce en las pequeñas arterias proximales.
La síntesis y la liberación de NO desde las células endoteliales están estimuladas asimismo por algunos vasoconstrictores, como la angiotensina II, que se unen a receptores específicos en las células endoteliales. El aumento en la liberación de NO protege contra un exceso de vasoconstricción. Cuando las células endoteliales resultan dañadas por hipertensión crónica o ateroesclerosis, la degradación en la síntesis de NO puede contribuir a una vasoconstricción excesiva y un empeoramiento de la hipertensión y a un daño endotelial, que, si no se trata, puede producir finalmente lesión vascular y daños en tejidos vulnerables como el corazón, los riñones y el encéfalo. Antes de que se descubriera el NO, los médicos utilizaban nitroglicerina, nitratos de amilo y otros derivados de nitratos para tratar a pacientes que sufrían angina de pecho, un dolor torácico intenso causado por isquemia del músculo cardíaco. Al descomponerse químicamente, estos fármacos liberan NO y provocan la dilatación de vasos sanguíneos de todo el organismo, incluidos los vasos sanguíneos coronarios. Otras aplicaciones importantes de la fisiología y la farmacología del NO son el desarrollo y el uso clínico de fármacos (p. ej., sildenafilo) que inhiben la fosfodiesterasa-5 (PDE-5) específica de cGMP, una enzima que degrada cGMP. Al impedir la degradación de cGMP, los inhibidores de PDE-5 prolongan eficazmente las acciones de NO para provocar vasodilatación. El uso clínico principal de los inhibidores de PDE-5 es el tratamiento de la disfunción eréctil.
La erección se produce por medio de impulsos de los nervios parasimpáticos a través de los nervios pélvicos en el pene, donde se liberan los neurotransmisores acetilcolina y NO. Al impedir la degradación de NO, los inhibidores de PDE-5 potencian la dilatación de los vasos sanguíneos en el pene y ayudan a la erección
Endotelina: un potente vasoconstrictor liberado por endotelio dañado.
Las células endoteliales también liberan sustancias vasoconstrictoras. La más importante de las mismas es la endotelina, un péptido de 21 aminoácidos que necesita sólo cantidades del orden de nanogramos para provocar una poderosa vasoconstricción. Esta sustancia está presente en las células endoteliales de todos o la mayoría de los vasos sanguíneos, aunque se eleva enormemente cuando los vasos resultan dañados. El estímulo habitual para la liberación es una lesión en el endotelio, como la provocada cuando se golpean los tejidos o se inyecta un producto químico traumatizante en el vaso sanguíneo. Después de un daño importante en los vasos sanguíneos, la liberación de endotelina local y la posterior vasoconstricción ayudan a evitar una hemorragia extensa de arterias de hasta 5 mm de diámetro que podrían haberse desgarrado por la lesión por aplastamiento. Se cree también que el aumento en la liberación de endotelina contribuye a la vasoconstricción cuando el endotelio sufre daños debidos a la hipertensión. Se han utilizado fármacos que bloquean los receptores de endotelina para tratar la hipertensión pulmonar, aunque en general no se han usado para reducir la presión arterial en pacientes con hipertensión arterial sistémica.
Regulación a largo plazo del flujo sanguíneo
Hasta este momento, la mayoría de los mecanismos de regulación del flujo sanguíneo local que hemos comentado actúan en pocos segundos o minutos después del cambio de la situación tisular local. A pesar de ello, el flujo sanguíneo se ajusta sólo en las tres cuartas partes de las necesidades adicionales de los tejidos, incluso después de la activación completa de estos mecanismos agudos. Por ejemplo, el flujo sanguíneo aumenta casi instantáneamente en un 100% cuando la presión arterial aumenta bruscamente desde 100 a 150 mmHg. En los 30 s a 2 min siguientes el flujo vuelve a disminuir hasta un 15% por encima del valor de control original, lo que demuestra la rapidez de los mecanismos agudos de regulación del flujo sanguíneo local pero, al mismo tiempo, se demuestra que la regulación aún es incompleta porque se mantiene un incremento del 15% del flujo sanguíneo. No obstante, en un período de horas, días o semanas, se desarrolla una regulación a largo plazo del flujo sanguíneo local que se suma al control agudo.
Esta regulación a largo plazo consigue un control mucho más completo del flujo sanguíneo. Por ejemplo, si la presión arterial se mantiene indefinidamente en 150 mmHg en el ejemplo anterior, en pocas semanas el flujo sanguíneo que atraviesa los tejidos se va aproximando gradualmente casi exactamente al nivel de flujo normal. . Obsérvese que una vez que la regulación a largo plazo ha tenido tiempo de desarrollarse, los cambios a largo plazo de la presión arterial entre 50 y 250 mmHg tienen poco efecto sobre la velocidad de flujo sanguíneo local. La regulación a largo plazo del flujo sanguíneo es especialmente importante cuando cambian las demandas metabólicas del tejido a largo plazo. Es decir, si un tejido está crónicamente hiperactivo y, por tanto, requiere un aumento crónico de las cantidades de oxígeno y otros nutrientes, por lo que en algunas semanas aumentan tanto el número como el tamaño de las arteriolas y los vasos capilares para cubrir las necesidades del tejido, a menos que el aparato circulatorio se vuelva patológico o sea demasiado viejo para responder.
Mecanismo de regulación a largo plazo: cambio de la «vascularización tisular»
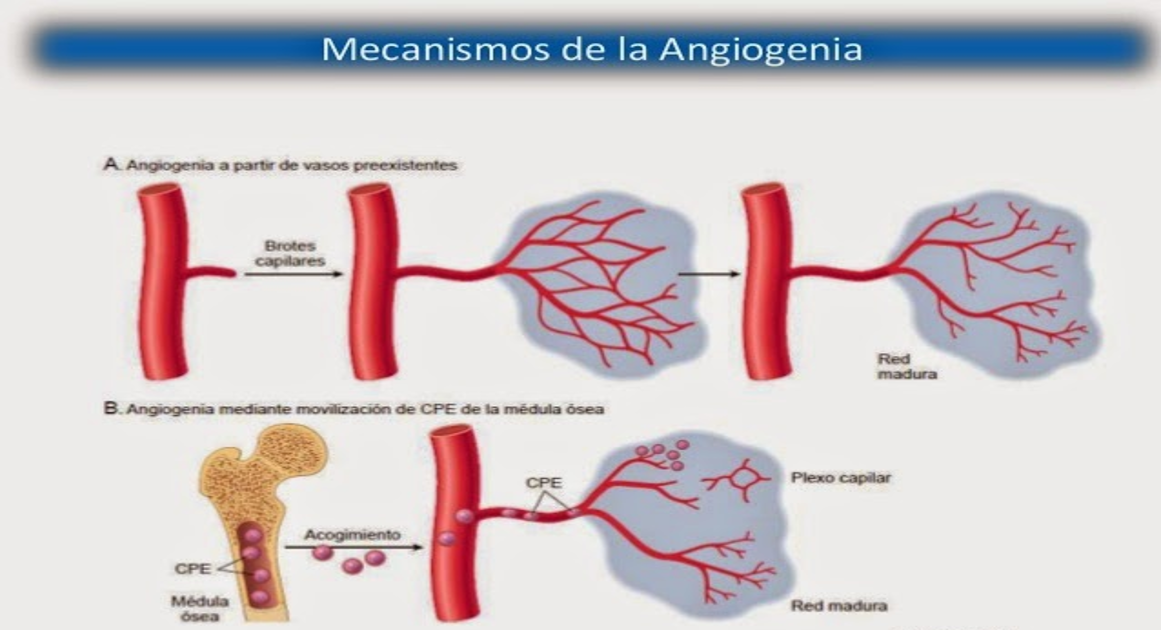
El mecanismo de regulación del flujo sanguíneo local a largo plazo consiste principalmente en cambiar la cantidad de vascularización de los tejidos. Por ejemplo, la vascularización aumenta si el metabolismo de un tejido dado aumenta durante un período prolongado, en un proceso denominado generalmente angiogenia; si el metabolismo disminuye, la vascularización también lo hace.
Es decir, se produce una reconstrucción física real de la vasculatura tisular para cubrirlas necesidades de los tejidos. Esta reconstrucción es rápida (en días) en los animales muy jóvenes y también en un tejido de nuevo crecimiento, como en el tejido cicatricial o el tejido canceroso, pero es más lenta en los tejidos antiguos y bien establecidos. Por tanto, el tiempo necesario para que tenga lugar la regulación a largo plazo puede ser de sólo unos días en el recién nacido o hasta meses en la tercera edad. Además, el grado último de respuesta es mucho mejor en tejidos más jóvenes que en los más mayores, por lo que la vascularización se ajustará en el recién nacido, para cubrir casi exactamente las necesidades de flujo sanguíneo del tejido, mientras que en los más antiguos la vascularización va por detrás de las necesidades de los tejidos.
Función del oxígeno en la regulación a largo plazo.
El oxígeno es importante no sólo para el control a corto plazo del flujo sanguíneo local, sino también para el control a largo plazo. Un ejemplo es el aumento de la vascularización de los tejidos en los animales que viven en altitudes elevadas, donde el oxígeno atmosférico es bajo. Un segundo ejemplo es que los fetos de pollo incubados con oxígeno bajo tienen hasta el doble de conductividad en el tejido de los vasos sanguíneos de lo normal. Este mismo efecto también se demuestra espectacularmente en los niños recién nacidos prematuros que se introducen en tiendas de oxígeno con fines terapéuticos. El exceso de oxígeno provoca la interrupción casi inmediata del crecimiento vascular nuevo en la retina de los ojos del niño prematuro e incluso provoca la degeneración de algunos de los vasos pequeños que ya se han formado. Después, cuando el niño es sacado de la tienda de oxígeno se produce un sobrecrecimiento explosivo de los vasos nuevos para compensar el descenso brusco del oxígeno disponible; en realidad, el sobrecrecimiento es tal que los vasos retinianos sobrepasan la retina hacia el humor vítreo del ojo, lo que terminará por provocar ceguera (afección que se conoce con el nombre de fibroplasia retrolental).
Importancia del factor de crecimiento endotelial vascular en la formación de vasos sanguíneos nuevos
Hay una docena o más de factores que aumentan el crecimiento de los vasos sanguíneos nuevos, siendo casi todos ellos péptidos pequeños. Tres de los mejor identificados son:
El factor de crecimiento de los fibroblastos
El factor de crecimiento del endotelio vascular (VEGF)
La angiogenina
Aislados cada uno de ellos en tejidos que tienen un aporte sanguíneo inadecuado. Presumiblemente, es la deficiencia de oxígeno tisular o de otros nutrientes la que provoca la formación de los factores de crecimiento vascular (también denominados «factores angiogénicos»). Prácticamente todos los factores angiogénicos favorecen el crecimiento de vasos nuevos del mismo modo, provocando la gemación desde otros vasos. El primer paso es la disolución de la membrana basal de las células endoteliales en el punto de gemación, seguida por la reproducción rápida de las células endoteliales nuevas que buscan la salida a través de la pared del vaso en cordones que se van extendiendo directamente hacia la fuente del factor angiogénico.
Las células de cada cordón continúan dividiéndose y se pliegan rápidamente formando un tubo. A continuación, este tubo se conecta con otro tubo que ha nacido de otro vaso donante (otra arteriola o vénula) y forma un asa capilar a través de la cual la sangre comienza a fluir. Si el flujo es suficientemente grande, los miocitos pequeños invaden finalmente la pared, por lo que algunos de los vasos nuevos finalmente se convertirán en arteriolas o vénulas nuevas o incluso en vasos más grandes. Es decir, la angiogenia explica la forma en que los factores metabólicos de los tejidos locales provocan el crecimiento de vasos nuevos. Algunas sustancias, como algunas hormonas esteroideas, tienen exactamente el efecto contrario sobre los vasos sanguíneos pequeños, en ocasiones causando incluso la disolución de las células vasculares y la desaparición de los vasos. Por tanto, los vasos sanguíneos también pueden desaparecer cuando no se necesitan. Los péptidos producidos en los tejidos pueden bloquear también el crecimiento de nuevos vasos sanguíneos. Por ejemplo, la angiostatina, un fragmento del plasminógeno proteico, es un inhibidor de la angiogenia de ocurrencia natural. La endostatina es otro péptido antiangiogénico que se deriva de la descomposición del colágeno tipo XVII. Aunque siguen sin conocerse las funciones fisiológicas precisas de estas sustancias antiangiogénicas, existe un gran interés en su uso potencial para detener el crecimiento de los vasos sanguíneos en tumores cancerosos y, por tanto, para prevenir los grandes aumentos en el flujo sanguíneo necesarios para sostener el suministro de nutrientes de tumores en rápido crecimiento.
La vascularización se encuentra determinada por la necesidad de flujo sanguíneo máximo, no por la necesidad media.
Una característica especial de gran valor del control vascular a largo plazo es que la vascularización se determina principalmente por el nivel máximo de flujo sanguíneo necesario y no por la necesidad media. Por ejemplo, durante el ejercicio intenso el flujo sanguíneo de todo el cuerpo aumenta el flujo sanguíneo en reposo hasta seis u ocho veces. Este mayor exceso de flujo puede no ser necesario más que durante algunos minutos cada día, aunque esta necesidad breve provoca la formación de VEGF suficiente en los músculos para aumentar su vascularización según necesidades. Si no fuera por esta capacidad, cada vez que una persona intentara hacer un ejercicio intenso los músculos no podrían recibir los nutrientes adecuados, en especial el oxígeno necesario, por lo que los músculos no se contraerían. No obstante, después del desarrollo de esta vascularización extra los vasos sanguíneos extra se mantienen contraídos, abriéndose para permitir el flujo extra sólo cuando existan estímulos locales apropiados, como la falta de oxígeno, los estímulos nerviosos vasodilatadores u otros estímulos que provoquen el flujo extra necesario.
Desarrollo de la circulación colateral: un fenómeno de regulación a largo plazo del flujo sanguíneo local
Cuando se bloquea una arteria o una vena en cualquier tejido del organismo se desarrolla un canal vascular nuevo rodeando el bloqueo y permitiendo que se vuelva a suministrar sangre al tejido afectado, al menos parcialmente. La primera etapa de este proceso es la dilatación de los bucles vasculares pequeños que ya conectan ese vaso proximal al bloqueo con el vaso distal. Esta dilatación se produce en el primer o segundo minutos, lo que indica que la dilatación está mediada probablemente por factores metabólicos que relajan las fibras musculares de los vasos pequeños implicados. Después de esta apertura inicial de los vasos colaterales, el flujo es menor de la cuarta parte de lo necesario para cubrir todas las necesidades tisulares. No obstante, la apertura se produce en las horas siguientes, por lo que antes de un día pueden estar ya cubiertas la mitad de las necesidades tisulares y en pocos días el flujo sanguíneo suele ser suficiente para cubrir todas estas necesidades. Los vasos colaterales continúan creciendo durante muchos meses después, casi siempre formando muchos canales colaterales pequeños en lugar de un único vaso de gran tamaño. En reposo, el flujo sanguíneo vuelve muy cerca de los valores normales, pero los nuevos canales son suficientemente grandes como para aportar el flujo sanguíneo necesario durante la actividad tisular agotadora. Es decir, el desarrollo de los vasos colaterales sigue los principios habituales del control a corto y largo plazo del flujo sanguíneo local, consistiendo el control a corto plazo en la dilatación metabólica rápida seguido crónicamente por el crecimiento e ingurgitación de los vasos nuevos en un período de semanas y meses.
El ejemplo más importante de desarrollo de los vasos sanguíneos colaterales lo encontramos después de la trombosis de una de las arterias coronarias. Casi todas las personas tienen cerrada a los 60 años al menos una rama menor de los vasos coronarios cerrados, o al menos parcialmente ocluidos. A pesar de ello, la mayoría de las personas no sabe que esto ha sucedido porque las colaterales se han desarrollado con la rapidez suficiente para prevenir el daño miocárdico. Es en los demás casos, en los que se desarrolla la insuficiencia coronaria con demasiada rapidez o con una intensidad excesiva para que se desarrollen las colaterales, cuando se desarrolla el ataque cardíaco.
Control humoral de la circulación
El control humoral de la circulación se refiere al control por las sustancias segregadas o absorbidas en los líquidos del organismo, como hormonas y factores producidos localmente. Algunas de esas sustancias se forman en glándulas especiales y se transportan en la sangre por todo el organismo, mientras que otras se forman en algunas zonas del tejido afectado y provocan sólo efectos circulatorios locales. Entre los factores humorales más importantes que afectan a la función circulatoria destacan los siguientes.
Sustancias vasoconstrictoras
Noradrenalina y adrenalina. La noradrenalina es una hormona vasoconstrictora especialmente potente; la adrenalina es menos potente y en algunos tejidos provoca incluso una vasodilatación leve. (Un ejemplo especial de vasodilatación provocada por la adrenalina es la dilatación coronaria durante el aumento de la actividad cardíaca.) Cuando se estimula el sistema nervioso simpático en el cuerpo durante el estrés o el ejercicio, las terminaciones nerviosas simpáticas de cada tejido liberan noradrenalina, que excita al corazón y contrae las venas y las arteriolas. Además, los nervios simpáticos de la médula suprarrenal provocan la secreción de noradrenalina y adrenalina en la sangre. Estas hormonas circulan entonces por todo el cuerpo y provocan casi los mismos efectos en la circulación que la estimulación simpática directa, con lo que se consigue un sistema de control doble:
1) estimulación nerviosa directa
2) efectos indirectos de la noradrenalina y/o de la adrenalina en la sangre circulante.
Angiotensina II. La angiotensina II es otra sustancia vasoconstrictora potente. Tan sólo una millonésima de gramo puede aumentar la presión arterial de un ser humano en 50 mmHg o más. El efecto de angiotensina II contrae potentemente las pequeñas arteriolas. Si esto sucede en un tejido aislado, el flujo sanguíneo de esa zona disminuirá mucho, aunque la importancia real de la angiotensina II es que normalmente actúa sobre muchas de las arteriolas del organismo al mismo tiempo, para aumentar la resistencia periférica total y aumentar la presión arterial. Es decir, esta hormona tiene un papel fundamental en la regulación de la presión arterial,
Vasopresina. La vasopresina, que también se conoce como hormona antidiurética, es aún más potente que la angiotensina II como vasoconstrictora, por lo que se convierte en una de las sustancias constrictoras más potentes del organismo. Se forma en las células nerviosas del hipotálamo, pero después se transporta distalmente a través de los axones nerviosos hacia la neurohipófisis, donde es finalmente segregada a la sangre. Es evidente que la vasopresina podría tener efectos muy importantes sobre la función circulatoria. Sin embargo, sólo se segrega en cantidades mínimas en condiciones normales, por lo que la mayoría de los fisiólogos opina que su papel en el control vascular es pequeño. No obstante, en estudios experimentales se ha demostrado que la concentración de vasopresina en sangre circulante puede aumentar después de una hemorragia intensa, lo suficiente como para elevar la presión arterial hasta en 60 mmHg. En muchos casos, este mecanismo puede elevar por sí solo la presión arterial hasta la normalidad. La vasopresina tiene una función importante aumentando la reabsorción de agua de los túbulos renales hacia la sangre, por tanto, ayudando a controlar el volumen de líquido corporal. De ahí viene el nombre de hormona antidiurética.
Sustancias vasodilatadoras
Bradicinina. Hay un grupo de sustancias denominadas cininas que provocan una vasodilatación potente cuando se forman en la sangre y en los líquidos tisulares de algunos órganos. Las cininas son pequeños polipéptidos que se escinden por enzimas proteolíticas a partir de 2-globulinas del plasma o los líquidos tisulares. Una enzima proteolítica de particular importancia para tal fin es la calicreína, que se encuentra en la sangre y los líquidos tisulares en una forma inactiva. Esta calicreína inactiva se activa por la maceración de la sangre, por la inflamación tisular o por otros efectos químicos o físicos similares. A medida que se va activando la calicreína actúa inmediatamente sobre la 2-globulina para liberar una cinina llamada calidina, que después se convierte en bradicinina gracias a las enzimas tisulares. Una vez formada, la bradicinina persiste durante sólo unos minutos, porque se inactiva por la enzima carboxipeptidasa o por la enzima convertidora, la misma que participa en la activación de la angiotensina. La enzima calicreína activada se destruye por un inhibidor de la calicreína que también está presente en los líquidos corporales. La bradicinina provoca una dilatación arteriolar potente y aumenta la permeabilidad capilar. Por ejemplo, la inyección de 1 mm de bradicinina en la arteria braquial de una persona aumenta el flujo sanguíneo a través del brazo hasta en seis veces, e incluso cantidades menores inyectadas localmente en los tejidos pueden provocar un edema local importante como consecuencia del aumento de tamaño de los poros capilares. Hay razones para creer que las cininas tienen un papel especial en la regulación del flujo sanguíneo y en la pérdida capilar de los líquidos en los tejidos inflamados. También parece que la bradicinina participa normalmente en la regulación del flujo sanguíneo en la piel y también en las glándulas salivares y gastrointestinales.
Histamina. La histamina se libera esencialmente en todos los tejidos del organismo cuando sufren daños o se inflaman, o cuando se sufre una reacción alérgica. La mayoría de la histamina deriva de los mastocitos en los tejidos dañados y de los basófilos en sangre. La histamina tiene un efecto vasodilatador potente sobre las arteriolas y, como la bradicinina, puede aumentar en gran medida la porosidad capilar permitiendo la pérdida tanto de líquidos como de proteínas plasmáticas hacia los tejidos. En muchas situaciones patológicas la dilatación arteriolar intensa y el aumento de la porosidad capilar producida por la histamina provoca la pérdida de cantidades enormes de líquido desde la circulación hacia los tejidos, induciendo el edema. Los efectos locales vasodilatadores y productores de edema de la histamina son especialmente prominentes durante las reacciones alérgicas
Control vascular por iones y otros factores químicos
Hay muchos iones y otros factores químicos que pueden dilatar o contraer los vasos sanguíneos locales. La mayoría de ellos tiene una función escasa en la regulación global de la circulación, pero hay algunos efectos específicos, como son:
El aumento de la concentración del ion calcio provoca vasoconstricción, que es consecuencia del efecto general del calcio para estimular la contracción del músculo liso,
El aumento de la concentración del ion potasio, dentro del intervalo fisiológico, provoca vasodilatación, que es consecuencia de la capacidad de los iones potasio para inhibir la contracción del músculo liso.
El aumento de la concentración del ion magnesio provoca una vasodilatación potente, porque los iones magnesio inhiben la contracción del músculo liso.
El aumento de la concentración del ion hidrógeno (descenso del pH) provoca la dilatación de las arteriolas. Por el contrario, un descenso pequeño de la concentración del ion hidrógeno provoca la constricción arteriolar.
Los aniones que tienen efectos significativos sobre los vasos sanguíneos son los iones acetato y citrato, que provocan una vasodilatación pequeña.
El aumento de la concentración de dióxido de carbono provoca una vasodilatación moderada en la mayoría de los tejidos, pero una vasodilatación importante en el cerebro. Además, el dióxido de carbono en la sangre tiene un efecto indirecto muy potente al actuar en el centro vasomotor del cerebro, transmitido a través del sistema nervioso simpático vasoconstrictor, provocando una vasoconstricción generalizada en todo el organismo. La mayoría de los vasodilatadores o vasoconstrictores tienen un efecto escaso en el flujo sanguíneo a largo plazo salvo que alteren la tasa metabólica de los tejidos. En la mayoría de los casos, el flujo sanguíneo en los tejidos y el gasto cardíaco (la suma del flujo en todos los tejidos del organismo) no se ven alterados sustancialmente, salvo durante uno o dos días, en estudios experimentales cuando se infunden crónicamente grandes cantidades de potentes vasoconstrictores como la angiotensina II o vasodilatadores como la bradicinina. ¿Por qué el flujo sanguíneo no se altera significativamente en la mayoría de los tejidos aun en presencia de cantidades muy elevadas de estos agentes vasoactivos? Para responder a esta pregunta debemos recordar uno de los principios fundamentales de la función circulatoria que hemos comentado anteriormente: la capacidad de cada tejido de autorregular su propio flujo sanguíneo de acuerdo con las necesidades metabólicas y otras funciones del mismo.
La administración de un potente vasoconstrictor, como la angiotensina II, puede provocar descensos transitorios en el flujo sanguíneo tisular y en el gasto cardíaco, aunque por lo común tiene un efecto escaso a largo plazo si no modifica la tasa metabólica de los tejidos. Análogamente, la mayo ría de los vasodilatadores provocan únicamente cambios a corto plazo en el flujo sanguíneo tisular y el gasto cardíaco si no alteran el metabolismo de los tejidos. Por tanto, el flujo sanguíneo está regulado generalmente de acuerdo con las necesidades específicas de los tejidos siempre y cuando la presión arterial sea adecuada para perfundir los tejidos.
Cuestionario
•1.-¿Cual mecanismo resalta la estrecha conexión existente entre la regulación del flujo sanguíneo local y el aporte de oxígeno y de otros nutrientes a los tejidos?. -La hiperemia reactiva
2.-¿Cual proceso metabolico ocurre cuando cualquier tejido se vuelve muy activo.? – La hiperemia activa
3.-¿Que funcion tiene el NO (Óxido nítrico) el los vasos sanguineos? -Es un vasodilatador que permite la relajacion de los vasos sanguineos
4.-¿Cual es la sustancia vasoconstrictora más importante de las células endoteliales? -Es la endotelina
5.- ¿Que sustancias son vasodilatadoras? -Bradicina y histamina
La función de la circulación consiste en atender las necesidades del organismo: transportar nutrientes hacia los tejidos del organismo, transportar los productos de desecho, transportar las hormonas de una parte del organismo . En general, mantener un entorno apropiado en todos los líquidos tisulares del organismo para lograr la supervivencia y funcionalidad de las células .
El corazón y los vasos sanguíneos están controlados, a su vez, de forma que proporcionan el gasto cardíaco y la presión arterial necesarios para garantizar el flujo sanguíneo necesario.
CARACTERÍSTICAS FÍSICAS DE LA CIRCULACIÓN
La circulación está divida en circulación sistémica y circulación pulmonar. Como la circulación sistémica aporta el flujo sanguíneo a todos los tejidos del organismo excepto los pulmones, también se conoce como circulación mayor o circulación periférica.
COMPONENTES FUNCIONALES DE LA CIRCULACIÓN
La función de las arterias consiste en transportar la sangre con una presión alta hacia los tejidos, motivo por el cual las arterias tienen unas paredes vasculares fuertes y unos flujos sanguíneos importantes con una velocidad alta.
Las arteriolas son las últimas ramas pequeñas del sistema arterial y actúan controlando los conductos a través de los cuales se libera la sangre en los capilares. Las arteriolas tienen paredes musculares fuertes que pueden cerrarlas por completo o que pueden, al relajarse, dilatar los vasos varias veces, con lo que pueden alterar mucho el flujo sanguíneo en cada lecho tisular en respuesta a sus necesidades.
La función de los capilares consiste en el intercambio de líquidos, nutrientes, electrólitos, hormonas y otras sustancias en la sangre y en el líquido intersticial. Para cumplir esta función, las paredes del capilar son muy finas y tienen muchos poros capilares diminutos, que son permeables al agua y a otras moléculas pequeñas.
Las vénulas recogen la sangre de los capilares y después se reúnen gradualmente formando venas de tamaño progresivamente mayor.
Las venas funcionan como conductos para el transporte de sangre que vuelve desde las vénulas al corazón; igualmente importante es que sirven como una reserva importante de sangre extra. Como la presión del sistema venoso es muy baja, las paredes de las venas son finas.
VOLÚMENES DE SANGRE EN LOS DISTINTOS COMPONENTES DE LA CIRCULACIÓN
Aproximadamente el 84% de todo el volumen de sangre del organismo se encuentra en la circulación sistémica y el 16% en el corazón y los pulmones. Del 84% que está en la circulación sistémica, el 64% está en las venas, el 13% en las arterias y el 7% en las arteriolas y capilares sistémicos. El corazón contiene el 7% de la sangre y los vasos pulmonares, el 9%. Resulta sorprendente el bajo volumen de sangre que hay en los capilares, aunque es allí donde se produce la función más importante de la circulación, la difusión de las sustancias que entran y salen entre la sangre y los tejidos.
SUPERFICIES TRANSVERSALES Y VELOCIDADES DEL FLUJO SANGUÍNEO
Si todos los vasos sistémicos de cada tipo se pusieran uno al lado del otro, la superficie transversal total aproximada para un ser humano medio sería la siguiente .La superficie transversal es mucho mayor de las venas que de las arterias, con una media cuatro veces mayor en las primeras, lo que explica la gran capacidad de reserva de sangre en el sistema venoso comparado con el sistema arterial. Como debe pasar el mismo volumen de flujo sanguíneo (F) a través de cada segmento de la circulación en cada minuto, la velocidad del flujo sanguíneo (v) es inversamente proporcional a la superficie transversal vascular (A). v = F/A Es decir, en condiciones de reposo la velocidad es como media de 33 cm/s en la aorta pero con una velocidad sólo de 1/1.000 en los capilares, es decir, aproximadamente 0,3 mm/s. No obstante, como los capilares tienen una longitud de sólo 0,3 a 1 mm, la sangre sólo se queda allí durante 13 s.
PRESIONES EN LAS DISTINTAS PORCIONES DE LA CIRCULACIÓN
El corazón bombea la sangre hacia la aorta, la presión media en este vaso es alta, con una media en torno a los 100 mmHg.El bombeo cardíaco es pulsátil, la presión arterial alterna entre una presión sistólica de 120 mmHg y una diastólica de 80 mmHg. A medida que el flujo sanguíneo atraviesa la circulación sistémica la presión media va cayendo progresivamente hasta llegar casi a 0 mmHg en el momento en el que alcanza la terminación de las venas cava, donde se vacía en la aurícula derecha del corazón. La presión de los capilares sistémicos oscila desde 35 mmHg cerca de los extremos arteriolares hasta tan sólo 10 mmHg cerca de los extremos venosos, pero la presión media «funcional» en la mayoría de los lechos vasculares es de 17 mmHg, una presión suficientemente baja que permite pequeñas fugas de plasma a través de los poros diminutos de las paredes capilares, aunque los nutrientes pueden difundir fácilmente a través de los mismos poros hacia las células de los tejidos externos.
En la parte derecha se ven las presiones en los distintos componentes de la circulación pulmonar. En las arterias pulmonares la presión es pulsátil, igual que en la aorta, pero la presión es bastante menor: la presión sistólica arterial pulmonar alcanza un promedio de 25 mmHg y la diastólica, de 8 mmHg, con una presión arterial pulmonar media de sólo 16 mmHg. La media de la presión capilar pulmonar alcanza un promedio de sólo 7 mmHg.El flujo sanguíneo por minuto a través de los pulmones es el mismo que en la circulación sistémica. Las bajas presiones del sistema pulmonar coinciden con las necesidades de los pulmones, lo único que se necesita es la exposición de la sangre en los capilares pulmonares al oxígeno y otros gases en los alvéolos pulmonares.
PRINCIPIOS BASICOS DE LA FUNCIÓN CIRCULATORIA
La velocidad del flujo sanguíneo en cada tejido del organismo casi siempre se controla con precisión en relación con la necesidad del tejido.
El gasto cardíaco se controla principalmente por la suma de todos los flujos tisulares locales.
La regulación de la presión arterial es generalmente independiente del control del flujo sanguíneo local o del control del gasto cardíaco.
papel importante en el control de la presión, tanto al se gregar hormonas que controlan la presión como al regular el volumen de sangre.
INTERRELACIONES ENTRE LA PRESIÓN,FLUJO Y RESISTENCIA
El flujo sanguíneo que atraviesa un vaso sanguíneo está determinado por dos factores: 1) diferencia de presión de la sangre entre los dos extremos de un vaso, también denominado «gradiente de presión» en el vaso, que es la fuerza que empuja la sangre a través del vaso, y 2) los impedimentos que el flujo sanguíneo encuentra en el vaso, que se conoce como resistencia vascular.
P1 representa la presión en el origen del vaso; en el otro extremo, la presión es P2. La resistencia es consecuencia de la fricción entre el flujo de sangre y el endotelio intravascular en todo el interior del vaso.
El flujo a través del vaso se puede calcular con la fórmula siguiente, que se conoce como ley de Ohm: F = ∆P/R en donde F es el flujo sanguíneo, ∆P es la diferencia de presión (P1 – P2) entre los dos extremos del vaso y R es la resistencia. En esta fórmula se afirma que el flujo sanguíneo es directamente proporcional a la diferencia de presión, pero inversamente proporcional a la resistencia.
FLUJO SANGUINEO
El flujo sanguíneo es la cantidad de sangre que atraviesa un punto dado de la circulación en un período de tiempo determinado.Se expresa en mililitros por minuto o litros por minuto, pero puede expresarse en mililitros por segundo o en cualquier otra unidad del flujo y de tiempo. El flujo sanguíneo global de toda la circulación de un adulto en reposo es de unos 5.000 ml/min, cantidad que se considera igual al gasto cardíaco porque es la cantidad de sangre que bombea el corazón en la aorta en cada minuto
MÉTODOS DE MEDICIÓN DEL FLUJO SANGUÍNEO
Se pueden introducir en serie muchos dispositivos mecánicos y electromecánicos dentro de un vaso sanguíneo o bien aplicarse en el exterior de la pared del vaso para medir el flujo. Este tipo de equipo se denomina flujómetro.
FLUJOMETRO ELECTROMAGNÉTICO
Uno de los dispositivos más importantes que permiten medir el flujo sanguíneo sin abrir el vaso es el flujómetro electromagnético, se muestra la generación de la fuerza electromotriz (voltaje eléctrico) de un cable que se mueve rápidamente atravesando un campo magnético. Este es el principio de producción de electricidad en un generador eléctrico,el mismo principio se aplica a la generación de una fuerza electromotriz en sangre que se está desplazando a través de un campo magnético. En este caso, se coloca un vaso sanguíneo entre los polos de un potente imán y se colocan los electrodos a ambos lados del vaso, perpendiculares a las líneas de fuerza magnéticas. Cuando el flujo sanguíneo atraviesa el vaso se genera entre los electrodos un voltaje eléctrico proporcional a la velocidad del flujo sanguíneo y el voltaje se registra usando un voltímetro o un aparato de registro electrónico apropiado. Una ventaja especial del flujómetro electromagnético es que puede registrar cambios del flujo en menos de 1/100 de segundo, con lo que se obtiene un registro exacto de los cambios pulsátiles del flujo y también de los valores en equilibrio.
FLUJÓMETRO ULTRASÓNICO DOPPLER
Se monta un cristal piezoeléctrico diminuto en el extremo de la pared del dispositivo. Cuando este cristal recibe la energía de un aparato eléctrico apropiado transmite una frecuencia de varios cientos de miles de ciclos por segundo distalmente sobre la sangre circulante. Una parte del sonido es reflejada por los eritrocitos de la sangre circulante y estas ondas de ultrasonidos reflejadas vuelven desde las células sanguíneas hacia el cristal con una frecuencia menor que la onda transmitida, porque los eritrocitos se están alejando del cristal transmisor. Es lo que se conoce como efecto Doppler , la onda de ultrasonido de alta frecuencia se interrumpe de forma intermitente y la onda reflejada vuelve hacia el cristal y se amplifica en el aparato electrónico, mientras que otra porción del mismo determina la diferencia de frecuencia entre la onda transmitida y la onda reflejada y, con ello, la velocidad del flujo sanguíneo. Siempre que no cambie el diámetro de un vaso sanguíneo, los cambios en el flujo sanguíneo en el vaso están relacionados directamente con las variaciones en la velocidad de flujo.El flujómetro ultrasónico Doppler puede registrar cambios pulsátiles .
FLUJO DE SANGRE LAMINAR EN LOS VASOS
Cuando el flujo sanguíneo se mantiene en equilibrio a través de un vaso sanguíneo largo y liso, el flujo se produce de forma aerodinámica, manteniéndose cada capa de sangre a la misma distancia de la pared del vaso. Además, la porción de sangre más central se mantiene en el centro del vaso. Este tipo de flujo se conoce como flujo laminar o flujo aerodinámico y es el contrario del flujo turbulento, que es el flujo sanguíneo que transcurre en todas las direcciones del vaso y se mezcla continuamente en su interior.
PERFIL DE VELOCIDAD PARABÓLICA DURANTE EL FLUJO LAMINAR
Cuando se produce el flujo laminar la velocidad del flujo en el centro del vaso es bastante mayor que la velocidad cerca de los bordes exteriores se muestra un vaso que contiene dos líquidos, el de la izquierda teñido con un colorante y el de la derecha, transparente, pero no hay flujo en el vaso. Cuando se hacen fluir, se desarrolla una interfase parabólica entre ellos, la porción de líquido adyacente a la pared del vaso apenas se ha movido, la que está algo alejada se ha desplazado una distancia pequeña y la que está en la porción central se ha desplazado mucho. Este efecto se conoce como el «perfil parabólico de la velocidad del flujo sanguíneo».
La causa de este perfil parabólico es la siguiente: las moléculas de líquido que tocan la pared se mueven lentamen te por su adherencia a la pared del vaso. La siguiente capa de moléculas se desliza sobre ellas, la tercera capa sobre la segunda, la cuarta sobre la tercera, etc. Por tanto, el líquido de la parte central del vaso se puede mover rápidamente porque hay muchas capas de moléculas deslizantes entre la zona central del vaso y su pared.
FLUJO DE SANGRE TURBULENTO EN ALGUNAS SITUACIONES
Cuando la velocidad del flujo sanguíneo es demasiado grande, cuando atraviesa una obstrucción en un vaso, hace un giro brusco o pasa sobre una superficie rugosa, el flujo puede volverse turbulento o desordenado en lugar de aerodinámico. El flujo turbulento significa que el flujo sanguíneo atraviesa el vaso en dirección transversal y también longitudinal, formando espirales que se denominan corrientes en torbellino, Cuando hay corrientes en torbellino el flujo sanguíneo encuentra una resistencia mucho mayor que cuando el flujo es aerodinámico, porque los torbellinos aumentan mucho la fricción global del flujo en el vaso. El flujo turbulento tiende a aumentar en proporción directa a la velocidad del flujo sanguíneo, al diámetro del vaso sanguíneo y a la densidad de la sangre y es inversamente proporcional a la viscosidad de la sangre.
Re = ν.d.ρ/n
Re es el número de Reynolds, una medida que da idea de la tendencia a producirse turbulencias, ( es la velocidad media del flujo sanguíneo (en centímetros/segundo), d es el diámetro del vaso (en centímetros), ρ es la densidad y n es la viscosidad (en poise). La viscosidad de la sangre suele ser de 1/30 poise y la densidad es sólo ligeramente mayor de 1, por lo que un aumento del número de Reynolds por encima de 200400 indica que se producirá flujo turbulento en algunas ramas de los vasos.
Condiciones apropiadas para que haya turbulencias:
1) una velocidad elevada del flujo sanguíneo
2) la naturaleza pulsátil del mismo
3) el cambio brusco del diámetro del vaso
4) un diámetro del vaso de gran calibre
PRESIÓN SANGUÍNEA
UNIDADES ESTÁNDAR DE PRESIÓN
La presión sanguínea se mide casi siempre en milímetros de mercurio (mmHg) porque el manómetro de mercurio se ha usado como patrón de referencia para medir la presión desde su invención en 1846 por Poiseuille. En realidad, la presión arterial mide la fuerza ejercida por la sangre contra una unidad de superficie de la pared del vaso.
Una presión de 1 mm de mercurio es igual a una presión de 1,36 cm de agua, porque la densidad del mercurio es 13,6 veces mayor que la del agua y 1 cm es 10 veces mayor que 1 mm.
RESISTENCIA AL FLUJO SANGUÍNEO
Unidades de resistencia.
La resistencia es el impedimento al flujo sanguíneo en un vaso, pero no se puede medir por medios directos. Por el contrario, la resistencia debe calcularse a partir de las determinaciones del flujo sanguíneo y de la diferencia de presión entre dos puntos del vaso. Si la diferencia de presión entre los dos puntos es de 1 mmHg y el flujo es de 1 ml/s, se dice que la resistencia es de una unidad de resistencia periférica, abreviada habitualmente como PRU.
EXPRESIÓN DE LA RESISTENCIA EN UNIDADES CGS
En ocasiones se usa una unidad física básica en CGS (centímetros, gramos, segundos) para expresar la resistencia. Esta unidad es la dina · s/cm5. La resistencia en esas unidades puede calcularse mediante la fórmula siguiente: R(en dina . s /cm5 ) = 1.333 × mmHg /ml/s
RESISTENCIA VASCULAR PERIFÉRICA TOTAL Y RESISTENCIA VASCULAR PULMONAR TOTAL
La velocidad del flujo sanguíneo a través de todo el sistema circulatorio es igual a la velocidad de la sangre que bombea el corazón. En un ser humano adulto es aproximadamente igual a 100 ml/s. La diferencia de presión entre las arterias sistémicas y las venas sistémicas es de unos 100 mmHg. Por tanto, la resistencia de toda la circulación sistémica, que se denomina resistencia periférica total, es de 100/100no 1 unidad de resistencia periférica PRU.
En el sistema pulmonar la presión arterial media es de 16 mmHg y la presión media en la aurícula izquierda es de 2 mmHg, con lo que la diferencia neta de presión es de 14 mm. Cuando el gasto cardíaco es normal, en torno a 100 ml/s, se calcula que la resistencia vascular pulmonar total es de 0,14 PRU (la séptima parte que en la circulación sistémica)
CONDUCTANCIA DE LA SANGRE EN UN VASO Y SU RELACIÓN CON LA RESISTENCIA
La conductancia es la medición del flujo sanguíneo a través de un vaso para dar una diferencia de presión dada. Se expresa en milímetros por segundo por milímetro de mercurio de presión, pero también se puede expresar en litros por segundo por milímetro de mercurio o en cualquier otra unidad del flujo sanguíneo y presión. Es evidente que la conductancia es el recíproco exacto de la resistencia según la ecuación: Conductancia = 1 /Resistencia
CAMBIOS MUY PEQUEÑOS EN EL DIÁMETRO DE UN VASO CAMBIAN MUCHO LA CONDUCTANCIA
Pequeños cambios en el diámetro de un vaso provocan cambios enormes en su capacidad de conducir la sangre cuando el flujo sanguíneo es aerodinámico.
LEY DE POISEUILLE
la sangre que está cerca de la pared de los vasos fluye lentamente, mientras que la que está en el centro del vaso fluye mucho más rápidamente. En el vaso pequeño, esencialmente toda la sangre está cerca de la pared, por lo que, sencillamente, no existe un chorro central de sangre que fluya con gran rapidez. Al integrar las velocidades de todos los anillos concéntricos de la sangre en movimiento y multiplicarlos por las superficies de los anillos se puede obtener la fórmula siguiente, que representa la ley de Poiseuille: F = π∆ Pr4 _ 81 en la que F es la velocidad del flujo sanguíneo, ∆P es la diferencia de presión entre los extremos del vaso, r es el radio del vaso, l es la longitud del vaso y es la viscosidad de la sangre.En esta ecuación la velocidad del flujo sanguíneo es directamente proporcional a la cuarta potencia del radio del vaso, por lo tanto diámetro de un vaso sanguíneo es el que tiene la mayor importancia entre todos estos factores para determinar la velocidad del flujo sanguíneo a través del vaso.
IMPORTANCIA DE LA LEY DE LA CUARTA POTENCIA DEL DIÁMETRO DEL VASO PARA DETERMINAR LA RESISTENCIA ARTERIOLAR
En la circulación sistémica, aproximadamente dos tercios de toda la resistencia sistémica al flujo sanguíneo se debe a la resistencia arteriolar en las pequeñas arteriolas. Los diámetros internos de las arteriolas varían desde tan sólo 4 mm hasta 25 mm,la ley de la cuarta potencia relaciona el vaso sanguíneo con el diámetro del vaso, se puede ver que este incremento en cuatro veces del diámetro del vaso aumenta el flujo hasta en 256 veces, es decir, esta ley de cuarta potencia hace que sea posible que las arteriolas, que responden con sólo pequeños cambios del diámetro a las señales nerviosas o a las señales químicas de los tejidos locales, hagan desaparecer casi completamente el flujo sanguíneo hacia el tejido o vayan al otro extremo, provocando un inmenso incremento del flujo.
RESISTENCIA AL FLUJO SANGUÍNEO EN CIRCUITO VASCULARES EN SERIE Y PARALELO
La sangre que bombea el corazón fluye desde la parte de presión alta de la circulación sistémica (es decir, la aorta) hacia el lado de baja presión a través de muchos miles de vasos sanguíneos dispuestos en serie y en paralelo. Las arterias, arteriolas, capilares, vénulas y venas se disponen colectivamente en serie. Cuando esto sucede, el flujo de cada vaso sanguíneo es el mismo y la resistencia total al flujo sanguíneo (Rtotal) es igual a la suma de la resistencia de cada vaso:
R total = R1 + R2 + R3 + R4
La resistencia vascular periférica total es igual a la suma de resistencias de las arterias, arteriolas, capilares, vénulas y venas.
Los vasos sanguíneos emiten numerosas ramas que forman circuitos paralelos que aportan la sangre a los distintos órganos y tejidos del organismo. Esta distribución paralela permite que cada tejido regule su propio flujo sanguíneo en mayor grado, independientemente del flujo de los demás tejidos. En cuanto a los vasos sanguíneos en paralelo, la resistencia total al flujo sanguíneo se expresa como: 1 _ Rtotal = 1 _ R1 + 1 _ R2 + 1 _ R3 + 1 _ R4 …
Es evidente que, para un gradiente de resistencia dado, fluirán cantidades de sangre mucho mayores a través de este sistema paralelo que a través de cada uno de los vasos sanguíneos por separado, por lo que la resistencia total es bastante menor que la resistencia de cualquier vaso sanguíneo aislado. El flujo a través de cada uno de los vasos unidos en paralelo de la figura 149B está determinado por el gradiente de presión y su propia resistencia, y no la resistencia de los demás vasos sanguíneos en paralelo. No obstante, el aumento de la resistencia de cualquiera de los vasos sanguíneos aumenta la resistencia vascular total. Si hay muchos vasos sanguíneos en paralelo será más sencillo para la sangre fluir a través del circuito porque cada vaso paralelo constituye otra vía o conductancia para el flujo sanguíneo.
La conductancia total (Ctotal) del flujo sanguíneo es la suma de la conductancia de cada vía paralela: C total = C1 + C2 + C3 + C4
EFECTO DEL HEMATOCRITO Y DE LA VISCOSIDAD DE LA SANGRE SOBRE LA RESISTENCIA VASCULAR Y EL FLUJO SANGUÍNEO
Otro de los factores importantes de la ley de Poiseuille es la viscosidad de la sangre. Cuanto mayor sea la viscosidad, menor será el flujo en un vaso si todos los demás factores se mantienen constantes. Además, la viscosidad de la sangre normal es tres veces mayor que la viscosidad del agua. Pero ¿qué hace que la sangre sea tan viscosa? Principalmente, el gran número de hematíes suspendidos en la sangre.
HEMATOCRITO
La proporción de la sangre que corresponde a glóbulos rojos se conoce como hematocrito, es decir, si una persona tiene un hematocrito de 40 significa que el 40% del volumen sanguíneo está formado por las células y el resto es plasma. El hematocrito de un varón adulto alcanza un promedio de 42, mientras que en las mujeres es de 38.
El hematocrito se determina centrifugando la sangre en el tubo calibrado.
EFECTO DEL HEMATOCRITO SOBRE LA VISCOSIDAD DE LA SANGRE
La viscosidad de la sangre aumenta drásticamente a medida que lo hace el hematocrito. La viscosidad de la sangre total con un hematocrito normal es de 3, lo que significa que se necesita tres veces más presión para obligar a la sangre total a atravesar un vaso que si fuera agua. Cuando el hematocrito aumenta hasta 60 o 70, como sucede en caso de policitemia, la viscosidad de la sangre puede ser hasta 10 veces mayor que la del agua y su flujo a través de los vasos sanguíneos se retrasa mucho. Otros factores que afectan a la viscosidad de la sangre son la concentración y el tipo de las proteínas plasmáticas. La viscosidad del plasma sanguíneo es 1,5 veces la del agua.
EFECTOS DE LA PRESIÓN SOBRE LA RESISTENCIA VASCULAR Y EL FLUJO SANGUÍNEO TISULAR
La «autorregulación» atenúa el efecto de la presión arterial en el flujo sanguíneo tisular. El incremento de la presión arterial debería provocar un incremento proporcional del flujo sanguíneo en los distintos tejidos del organismo. La razón de este incremento es que el aumento de la presión arterial no sólo aumenta la fuerza que impulsa la sangre a través de los vasos, sino que también inicia incrementos compensatorios en la resistencia vascular a través de la activación de los mecanismos locales de control. De modo inverso, con las reducciones en la presión arterial, la mayor parte de la resistencia vascular se reduce en un tiempo breve en los tejidos y el flujo sanguíneo se mantiene relativamente constante. La capacidad de cada tejido de ajustar su resistencia vascular y mantener un flujo sanguíneo normal durante los cambios en la presión arterial entre aproximadamente 70 y 175 mmHg se denomina AUTORREGULACIÓN DEL FLUJO SANGUÍNEO.
Los cambios del flujo sanguíneo se pueden provocar mediante la estimulación simpática, que constriñe los vasos sanguíneos periféricos. Análogamente, los vasoconstrictores hormonales, como noradrenalina, angiotensina II, vasopresina o endotelina, también pueden reducir el flujo sanguíneo.
RELACIÓN PRESIÓN-FLUJO EN LOS LECHOS VASCULARES PASIVOS
En vasos sanguíneos aislados o en tejidos que no muestran autorregulación, los cambios en la presión arterial pueden tener efectos importantes en el flujo sanguíneo. El motivo es que el aumento de la presión arterial no sólo incrementa la fuerza que impulsa la sangre a través de los vasos sino que además distiende los vasos elásticos, para reducir en la práctica la resistencia vascular.
Inversamente, el descenso en la presión arterial en vasos sanguíneos pasivos eleva la resistencia, ya que los vasos elásticos se colapsan gradualmente debido a la reducción en la presión de distensión. Cuando la presión desciende por debajo de un nivel crítico, denominado presión de cierre crítica, el flujo cesa en el momento en que los vasos sanguíneos se colapsan por completo. La estimulación simpática y otros vasoconstrictores pueden alterar la relación de flujopresión.
la inhibición de la actividad simpática dilata mucho los vasos y aumenta el flujo sanguíneo al doble o más. Por el contrario, una estimulación simpática potente contrae los vasos tanto que, en ocasiones, el flujo sanguíneo disminuye casi a cero durante unos segundos
El corazón está dotado de un sistema especial para:
1) generar impulsos eléctricos rítmicos para producir la contracción rítmica del músculo cardíaco.
2) conducir estos estímulos rápidamente por todo el corazón.
Cuando este sistema funciona normalmente, las aurículas se contraen aproximadamente 1/6 de segundo antes de la contracción ventricular, lo que permite el llenado de los ventrículos antes de que bombeen la sangre a través de los pulmones y de la circulación periférica.
Este sistema rítmico y de conducción del corazón se puede lesionar en las cardiopatías, especialmente en la isquemia de los tejidos cardíacos que se debe a una disminución del flujo sanguíneo coronario. La consecuencia es con frecuencia una alteración del ritmo cardíaco o una secuencia anormal de contracción de las cavidades cardíacas.
SISTEMA DE EXCITACIÓN ESPECIALIZADO Y DE CONDUCCIÓN DEL CORAZÓN
En el sistema de excitación tendremos e nódulo sinusal (también denominado nódulo sinoauricular o SA), en el que se genera el impulso rítmico normal; las vías internodulares que conducen impulsos desde el nódulo sinusal hasta el nódulo auriculoventricular (AV); el nódulo AV, en el cual los impulsos originados en las aurículas se retrasan antes de penetrar en los ventrículos; el haz AV, que conduce impulsos desde las aurículas hacia los ventrículos, y las ramas izquierda y derecha del haz de fibras de Purkinje, que conducen los impulsos cardíacos por todo el tejido de los ventrículos.
NÓDULO SINUSAL (SINOAURICULAR)
El nódulo sinusal (también denominado nódulo sinoauricular) es una banda elipsoide, aplanada y pequeña de músculo cardíaco especializado de aproximadamente 3 mm de anchura, 15 mm de longitud y 1 mm de grosor. Está localizado en la pared posterolateral superior de la aurícula derecha, inmediatamente inferior y ligeramente lateral a la desembocadura de la vena cava superior.
las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares, de modo que todos los potenciales de acción que comienzan en el nódulo sinusal se propagan inmediatamente hacia la pared del músculo auricular.
RITMICIDAD ELÉCTRICA AUTOMÁTICA DE LAS FIBRAS SINUSALES
Algunas fibras cardíacas tiene la capacidad de autoexcitación, que es un proceso que puede producir descargas y contracciones rítmicas automáticas. Esto es especialmente cierto en el caso de las fibras del sistema especializado de conducción del corazón, entre ellas las fibras del nódulo sinusal. Por este motivo el nódulo sinusal habitualmente controla la frecuencia del latido de todo el corazón.
MECANISMOS DE LA RITMICIDAD DEL NÓDULO SINUSAL
muestra potenciales de acción registrados desde el interior de una fibra del nódulo sinusal durante tres latidos cardíacos y, a modo de comparación, un único potencial de acción de una fibra muscular ventricular.Entre descargas el «potencial de membrana en reposo» de la fibra del nódulo sinusal tiene una negatividad de aproximadamente –55 a –60 mV, en comparación con –85 a –90 mV para la fibra muscular ventricular. La causa de esta menor negatividad es que las membranas celulares de las fibras sinusales son permeables naturalmente a los iones sodio y calcio, y las cargas positivas de los iones sodio y calcio que entran neutralizan parte de la negatividad intracelular.
El músculo cardíaco tiene tres tipos de canales iónicos de membrana que tienen funciones importantes en la generación de los cambios de voltaje en el potencial de acción.
Los tipos son:
1) los canales rápidos de sodio
2) los canales lentos de sodio-calcio
3) los canales de potasio.
La apertura de los canales rápidos de sodio es responsable de la rápida espiga ascendente del potencial de acción que se observa en el músculo ventricular, debido a la entrada rápida de iones sodio positivos hacia el interior de la fibra. Después, la «meseta» del potencial de acción ventricular está producida por la apertura lenta de los canales lentos de sodiocalcio, que dura aproximadamente 0,3 s. Finalmente, la apertura de los canales de potasio permite la difusión de grandes cantidades de iones potasio positivos hacia el exterior a través de la membrana de la fibra y devuelve el potencial de membrana a su nivel de reposo.
Hay una diferencia en la función de estos canales en la fibra del nódulo sinusal porque el potencial «de reposo» es mucho menos negativo (de sólo –55 mV en la fibra nodular, en lugar de los –90 mV de la fibra muscular ventricular). A este nivel de –55 mV, los canales rápidos de sodio principalmente ya se han «inactivado», lo que significa que han sido bloqueados. La causa de esto es que siempre que el potencial de membrana es menos negativo –55 mV durante más de algunos milisegundos, las compuertas inactivación del interior de la membrana celular que cierran los canales rápidos de sodio se cierran y permanecen de esta manera. Por tanto, sólo se pueden abrir los canales lentos de sodiocalcio (es decir, se pueden «activar») y, por tanto, pueden producir el potencial de acción. En consecuencia, el potencial de acción del nódulo auricular se produce más lentamente que el potencial de acción del músculo ventricular
AUTOEXCITACIÓN DE LAS FIBRAS DEL NÓDULO SINUSAL
Debido a la elevada concentración de iones sodio en el líquido extracelular en el exterior de la fibra nodular, así como al número moderado de canales de sodio abiertos previamente, los iones sodio positivos del exterior de las fibras normalmente tienden a desplazarse hacia el interior. Por tanto, entre los latidos cardíacos, la entrada de iones sodio de carga positiva produce una elevación lenta del potencial de membrana en reposo en dirección positiva. El potencial «en reposo» aumenta gradualmente y se hace menos negativo entre cada dos latidos sucesivos. Cuando el potencial alcanza un voltaje umbral de aproximadamente –40 mV, los canales de sodiocalcio se «activan», produciendo de esta manera el potencial de acción.
La permeabilidad inherente de las fibras del nódulo sinusal a los iones sodio y calcio produce su autoexcitación.
El estado de hiperpolarización inicialmente desplaza el potencial de membrana «en reposo» hacia abajo hasta aproximadamente –55 o –60 mV al final del potencial de acción. ¿Por qué este nuevo estado de hiperpolarización no se mantiene indefinidamente? El motivo es que en las décimas de segundo siguientes al final del potencial de acción se cierran cada vez más canales de potasio. Los iones sodio y calcio que fluyen hacia el interior una vez más compensan el flujo de salida de iones potasio, y esto hace que el potencial «de reposo» se desplace hacia arriba una vez más, alcanzando finalmente el nivel liminal de aproximadamente –40 mV. Después comienza de nuevo todo el proceso: autoexcitación .
VÍAS INTERNODULARES Y TRANSMISIÓN DEL IMPULSO CARDÍACO A TRAVÉS DE LAS AURÍCULAS
Los extremos de las fibras del nódulo sinusal se conectan directamente con las fibras musculares auriculares circundantes. Por tanto, los potenciales de acción que se originan en el nódulo sinusal viajan hacia estas fibras musculares auriculares. De esta manera, el potencial de acción se propaga por toda la masa muscular auricular y, finalmente, llega hasta el nódulo AV.
La velocidad de conducción en la mayor parte del músculo auricular es de aproximadamente 0,3 m/s, pero la conducción es más rápida, de aproximadamente 1 m/s, en varias pequeñas bandas de fibras auriculares. Una de ellas, denominada banda interauricular anterior, atraviesa las paredes anteriores de las aurículas para dirigirse hacia la aurícula izquierda. Además, otras tres bandas pequeñas se incurvan a través de las paredes auriculares anterior, lateral y posterior, y terminan en el nódulo AV; se denominan, vías internodulares anterior, media y posterior. La causa de la velocidad de conducción más rápida de estas bandas es la presencia de fibras de conducción especializadas.
NÓDULO AURICULOVENTRICULAR Y RETRASO DE LA CONDUCCIÓN DEL IMPULSO DESDE LAS AURICULAS A LOS VENTRICULOS
El sistema de conducción auricular está organizado de modo que el impulso cardíaco no viaja desde las aurículas hacia los ventrículos demasiado rápidamente; este retraso da tiempo para que las aurículas vacíen su sangre hacia los ventrículos antes de que comience la contracción ventricular.
El retraso de la transmisión hacia los ventrículos se produce principalmente en el nódulo AV y en sus fibras de conducción adyacentes. El nódulo AV está localizado en la pared posterolateral de la aurícula derecha, inmediatamente detrás de la válvula tricúspide.
Obsérvese que el impulso, después de viajar por las vías internodulares, llega al nódulo AV aproximadamente 0,03 s después de su origen en el nódulo sinusal. Después hay un retraso de otros 0,09 s en el propio nódulo AV antes de que el impulso entre en la porción penetrante del haz AV, a través del cual pasa hacia los ventrículos. Se produce un retraso final de otros 0,04 s principalmente en este haz AV penetrante, que está formado por múltiples fascículos pequeños que atraviesan el tejido fibroso que separa las aurículas de los ventrículos.
Así, el retraso total en el nódulo AV y en el sistema de AV es de aproximadamente 0,13 s. Esto, añadido al retraso inicial de la conducción de 0,03 s desde el nódulo sinusal hasta el nódulo AV, hace que haya un retraso total de 0,16 s antes de que la señal excitadora llegue finalmente al músculo ventricular que se está contrayendo.
CAUSA DE LA CONDUCCIÓN LENTA
La conducción lenta en las fibras transicionales, nodulares y penetrantes del haz AV está producida principalmente por la disminución del número de uniones en hendidura entre células sucesivas de las vías de conducción, de modo que hay una gran resistencia a la conducción de los iones excitadores desde una fibra de conducción hasta la siguiente. Por tanto, es fácil ver por qué se tarda en excitar células sucesivas.
TRANSMISIÓN RÁPIDA EN EL SISTEMA DE PURKINJE VENTRICULAR
Las fibras de Purkinje especiales se dirigen desde el nódulo AV a través del haz AV hacia los ventrículos. Excepto en la porción inicial de estas fibras, donde penetran en la barrera fibrosa AV, tienen características funcionales bastante distintas a las de las fibras del nódulo AV. Son fibras muy grandes, incluso mayores que las fibras musculares ventriculares normales, y transmiten potenciales de acción a una velocidad de 1,5 a 4,0 m/s, una velocidad aproximadamente 6 veces mayor que la del músculo ventricular normal y 150 veces mayor que la de algunas de las fibras del nódulo AV. Esto permite una transmisión casi instantánea del impulso cardíaco por todo el resto del músculo ventricular.
CONDUCCIÓN UNIDIRECCIONAL A TRAVÉS DEL HAZ AV
Una característica especial del haz AV es la imposibilidad, excepto en estados anormales, de que los potenciales de acción viajen retrógradamente desde los ventrículos hacia las aurículas. Esto impide la reentrada de los impulsos cardíacos por esta ruta desde los ventrículos hacia las aurículas, permitiendo sólo la contracción anterógrada desde las aurículas hacia los ventrículos. Además, se debe recordar que en todas las localizaciones excepto en el haz AV el músculo auricular está separado del músculo ventricular por una barrera fibrosa continua. Esta barrera normalmente actúa como aislante para impedir el paso de los impulsos cardíacos entre el músculo auricular y ventricular a través de cualquier ruta distinta a la conducción anterógrada a través del propio haz AV.
DISTRIBUCIÓN DE LAS FIBRAS DE PURKINJE EN LOS VENTRICULOS:LAS RAMAS IZQUIERDA Y DERECHA DEL HAZ
Después de penetrar en el tejido fibroso que está entre el músculo auricular y ventricular, la porción distal del haz AV se dirigía hacia abajo en el interior del tabique interventricular a lo largo de 5 a 15 mm hacia la punta del corazón. Después el haz se divide en las ramas izquierda y derecha del haz, que están debajo del endocardio en los dos lados respectivos del tabique interventricular. Cada una de las ramas se dirige hacia abajo, hacia la punta del ventrículo, dividiéndose progresivamente en ramas más pequeñas. Estas ramas, a su vez, siguen un trayecto en dirección lateral alrededor de cada una de las cavidades ventriculares y hacia atrás, hacia la base del corazón. Los extremos de las fibras de Purkinje penetran en aproximadamente un tercio del grosor de la masa muscular y finalmente se continúan con las fibras musculares cardíacas.
Desde el momento en el que el impulso cardíaco entre las ramas del haz en el tabique interventricular hasta que sale de las terminaciones de las fibras de Purkinje el tiempo total transcurrido es en promedio de sólo 0,03 s. Por tanto, una vez que el impulso cardíaco ha entrado en el sistema de conducción ventricular de Purkinje, se propaga casi inmediatamente a toda la masa del músculo ventricular.
TRANSMISIÓN DEL IMPULSO CARDÍACO EN EL MÚSCULO VENTRICULAR
Una vez que el impulso llega a los extremos de las fibras de Purkinje se transmite a través de la masa del músculo ventricular por las propias fibras musculares ventriculares. La velocidad de transmisión es ahora sólo de 0,3 a 0,5 m/s, una sexta parte de la velocidad de las fibras de Purkinje. El músculo cardíaco envuelve el corazón en una doble espiral, con tabiques fibrosos entre las capas en espiral; por tanto, el impulso cardíaco no viaja necesariamente directamente hacia el exterior, hacia la superficie del corazón, sino que se angula hacia la Superficie a lo largo de las direcciones de las espirales. Debido a esto, la transmisión desde la superficie endocárdica a la superficie epicárdica del ventrículo precisa hasta otros 0,03 s.El tiempo total para la transmisión del impulso cardíaco desde las ramas iniciales del haz hasta las últimas fibras del músculo ventricular en el corazón normal es de aproximadamente 0,06 s.
RESUMEN DE LA PROPAGACIÓN DEL IMPULSO CARDÍACO A TRAVÉS DEL CORAZÓN
El impulso se propaga a una velocidad moderada a través de las aurículas, aunque se retrasa más de 0,1 s en la región del nódulo AV antes de aparecer en el haz AV del tabique interventricular. Una vez que ha entrado en este haz, se propaga muy rápidamente a través de las fibras de Purkinje por todas las superficies endocárdicas de los ventrículos. Después el impulso se propaga de nuevo algo más lentamente a través del músculo ventricular hacia las superficies epicárdicas.
CONTROL DE LA EXCITACIÓN Y LA CONDUCCIÓN EN EL CORAZÓN
NÓDULO SINUSAL COMO MARCAPASOS DEL CORAZÓN
El impulso normalmente se origina en el nódulo sinusal. En algunas situaciones anormales no ocurre así. Otras partes del corazón pueden presentar también una excitación rítmica intrínseca de la misma forma que lo hacen las fibras del nódulo sinusal; esto es particularmente cierto en el caso de las fibras del nódulo AV y de las fibras de Purkinje. Las fibras del nódulo AV, cuando no son estimuladas por alguna fuente externa, descargan a una frecuencia rítmica intrínseca de 40 a 60 veces por minuto, y las fibras de Purkinje lo hacen a una frecuencia de entre 15 y 40 veces por minuto. Estas frecuencias son distintas a la frecuencia normal del nódulo sinusal, de 70 a 80 veces por minuto.
¿Por qué entonces es el nódulo sinusal, y no el nódulo AV ni las fibras de Purkinje, el que controla la ritmicidad del corazón? La frecuencia de descarga del nódulo sinusal es mayor que la frecuencia de descarga autoexcitadora natural de las fibras del nódulo AV y de las fibras de Purkinje. Cada vez que se produce una descarga en el nódulo sinusal su impulso se conduce hacia el nódulo AV y hacia las fibras de Purkinje, produciendo también la descarga de sus membranas. Pero el nódulo sinusal produce una nueva descarga antes de que las fibras del nódulo AV o las fibras de Purkinje puedan alcanzar sus propios umbrales de autoexcitación. Por tanto, el nuevo impulso procedente del nódulo sinusal descarga tanto las fibras del nódulo AV como las fibras de Purkinje antes de que se pueda producir autoexcitación en cualquiera de esas estructuras. El el nódulo sinusal controla el latido del corazón porque su frecuencia de descarga rítmica es más rápida que la de cualquier otra parte del corazón.
IMPORTANCIA DEL SISTEMA DE PURKINJE EN LA GENERACIÓN DE LA CONTRACCIÓN SINCRÓNICA DEL MÚSCULO VENTRICULAR
Normalmente el impulso cardíaco llega a casi todas las porciones de los ventrículos en un breve intervalo de tiempo, excitando la primera fibra muscular ventricular sólo 0,03 a 0,06 s antes de la excitación de la última. Esto hace que todas las porciones del músculo de los dos ventrículos comiencen a contraerse casi al mismo tiempo y que después sigan contrayéndose durante aproximadamente otros 0,3 s. La función de bomba eficaz de las dos cavidades ventriculares precisa este tipo sincrónico de contracción. Si el impulso cardíaco viajara lentamente a través de los ventrículos, buena parte de la masa ventricular se contraería antes de la contracción del resto, en cuyo caso se produciría una gran disminución de la función global de bomba. De hecho, en algunos tipos de trastornos cardíacos,se produce una transmisión lenta, y la eficacia del bombeo de los ventrículos disminuye hasta el 20-30%.
CONTROL DEL RITMO CARDÍACO Y LA CONDUCCIÓN DE IMPULSOS POR LOS NERVIOS CARDÍACOS: LOS NERVIOS SIMPÁTICOS Y PARASIMPÁTICOS
El corazón está inervado por nervios simpáticos y parasimpáticos, Los nervios parasimpáticos (vagos) se distribuyen principalmente a los nódulos SA y AV, en mucho menor grado al músculo de las dos aurículas y apenas directamente al músculo ventricular. Por el contrario, los nervios simpáticos se distribuyen en todas las regiones del corazón, con una intensa representación en el músculo ventricular, así como en todas las demás zonas.
LA ESTIMULACIÓN PARASIMPÁTICA (VAGAL) PUEDE RETRASAR O BLOQUEAR EL RITMO Y LA CONDUCCIÓN CARDIACA( ESCAPE VENTRICULAR)
La estimulación de los nervios parasimpáticos que llegan al corazón (los vagos) hace que se libere la hormona acetilcolina en las terminaciones nerviosas. Esta hormona tiene dos efectos principales sobre el corazón. Primero, reduce la frecuencia del ritmo del nódulo sinusal, y segundo, reduce la excitabilidad de las fibras de la unión AV entre la musculatura auricular y el nódulo AV, retrasando de esta manera la transmisión del impulso cardíaco hacia los ventrículos.
Una estimulación vagal débil a moderada reduce la frecuencia del bombeo del corazón, con frecuencia hasta un valor tan bajo como la mitad de lo normal. La estimulación intensa de los nervios vagos puede interrumpir completamente la excitación rítmica del nódulo sinusal o puede bloquear completamente la transmisión del impulso cardíaco desde las aurículas hacia los ventrículos a través del nódulo AV. Los ventrículos dejan de latir durante 5 a 20 s, pero después algún área pequeña de las fibras de Purkinje, habitualmente en la porción del tabique interventricular del haz AV, presenta un ritmo propio y genera la contracción ventricular a una frecuencia de 15 a 40 latidos por minuto. Este fenómeno se denomina escape ventricular.
MECANISMOS DE LOS EFECTOS VAGALES
La acetilcolina que se libera en las terminaciones nerviosas vagales aumenta mucho la permeabilidad de las membranas de las fibras a los iones potasio, lo que permite la salida rápida de potasio desde las fibras del sistema de conducción. Esto da lugar a un aumento de la negatividad en el interior de las fibras, un efecto que se denomina hiperpolarización, que hace que este tejido excitable sea mucho menos excitable. En el nódulo sinusal, el estado de hiperpolarización reduce el potencial de membrana «en reposo» de las fibras del nódulo sinusal a un nivel mucho más negativo de lo habitual, hasta –65 a –75 mV en lugar del nivel normal de –55 a –60 mV.
Por tanto, el aumento inicial del potencial de membrana del nódulo sinusal que produce la corriente de entrada de sodio y de calcio tarda mucho más en alcanzar el potencial liminal para la excitación. Esto retrasa mucho la frecuencia de ritmicidad de estas fibras nodulares. Si la estimulación vagal es lo suficientemente intensa es posible detener totalmente la autoexcitación rítmica de este nódulo. En el nódulo AV, el estado de hiperpolarización producido por la estimulación vagal hace que sea difícil que las pequeñas fibras auriculares que entran en el nódulo generen una corriente de una intensidad suficiente como para excitar las fibras nodulares. Por tanto, el factor de seguridad para la transmisión del impulso cardíaco a través de las fibras de transición hacia las fibras del nódulo AV disminuye.
EFECTO DE LA ESTIMULACIÓN SIMPÁTICA SOBRE EL RITMO Y LA CONDUCCIÓN DEL CORAZÓN
La estimulación simpática produce esencialmente los efectos contrarios sobre el corazón a los que produce la estimulación vagal.Primero, aumenta la frecuencia de descarga del nódulo sinusal. Segundo, aumenta la velocidad de conducción, así como el nivel de excitabilidad de todas las porciones del corazón. Tercero, aumenta mucho la fuerza de contracción de toda la musculatura cardíaca, tanto auricular como ventricular.La estimulación simpática aumenta la actividad global del corazón. La estimulación máxima casi puede triplicar la frecuencia del latido cardíaco y puede aumentar la fuerza de la contracción del corazón hasta dos veces.
MECANISMO DEL EFECTO SIMPÁTICO
La estimulación de los nervios simpáticos libera la hormona noradrenalina en las terminaciones nerviosas simpáticas. La noradrenalina estimula, a su vez, los receptores b1-adrenérgicos, que median en los efectos sobre la frecuencia cardíaca, aumenta la permeabilidad de la membrana de las fibras a los iones sodio y calcio. En el nódulo sinusal, un aumento de la permeabilidad a sodio-calcio genera un potencial en reposo más positivo y también produce un aumento de la velocidad del ascenso del potencial de membrana diastólico hacia el nivel liminal para la autoexcitación, acelerando de esta forma la autoexcitación y, por tanto, aumentando la frecuencia cardíaca.
En el nódulo AV y en los haces AV, el aumento de la permeabilidad a sodiocalcio hace que sea más fácil que el potencial de acción excite todas las porciones sucesivas de los haces de las fibras de conducción, disminuyendo de esta manera el tiempo de conducción desde las aurículas hasta los ventrículos. El aumento de la permeabilidad a los iones calcio es responsable al menos en parte del aumento de la fuerza contráctil del músculo cardíaco bajo la influencia de la estimulación simpática, porque los iones calcio tienen una función importante en la excitación del proceso contráctil de las miofibrillas.
El corazón está formado realmente por dos bombas separadas: un corazón derecho que bombea sangre hacia los pulmones y un corazón izquierdo que bombea sangre a través de la circulación sistémica que aporta flujo sanguíneo a los demás órganos y tejidos del cuerpo. A su vez, cada uno de estos corazones es una bomba bicameral pulsátil formada por una aurícula y un ventrículo. Cada una de las aurículas es una bomba débil de cebado del ventrículo, que contribuye a transportar sangre hacia el ventrículo correspondiente.
Los ventrículos después aportan la principal fuerza del bombeo que impulsa la sangre:
1) hacia la circulación pulmonar por el ventrículo derecho, o 2) hacia la circulación sistémica por el ventrículo izquierdo.
Mecanismos especiales del corazón producen una sucesión continuada de contracciones cardíacas denominada ritmicidad cardíaca, que transmite potenciales de acción por todo el músculo cardíaco y determina su latido rítmico.
El corazón está formado por tres tipos principales de músculo cardíaco: músculo auricular, músculo ventricular y fibras musculares especializadas de excitación y de conducción. El músculo auricular y ventricular se contrae de manera muy similar al músculo esquelético, excepto porque la duración de la contracción es mucho mayor. No obstante, las fibras especializadas de excitación y de conducción del corazón se contraen solo débilmente porque contienen pocas fibrillas contráctiles; en cambio, presentan descargas eléctricas rítmicas automáticas en forma de potenciales de acción o conducción de los potenciales de acción por todo el corazón, formando así un sistema excitador que controla el latido rítmico cardíaco.
ANATOMÍA FISIOLÓGICA DEL MÚSCULO CARDÍACO
La
histología del músculo cardíaco, que presenta las fibras musculares cardíacas dispuestas
en un retículo,
de modo que las fibras se dividen, se vuelven a combinar y se separan de nuevo.
Cabe observar que el músculo cardíaco es estriado,
igual que el músculo esquelético. Además, el músculo cardíaco tiene las
miofibrillas típicas que contienen
filamentos de actina y de miosina casi idénticos
a los que se encuentran en el músculo esquelético; estos filamentos están unos
al lado de otros y se deslizan durante la contracción de la misma manera que
ocurre en el músculo esquelético.
EL MÚSCULO CARDÍACO ES UN SINCITIO
Las
zonas oscuras que atraviesan las fibras musculares cardíacas se denominan discos
intercalados; realmente son membranas
celulares que separan las células musculares cardíacas individuales entre sí.
Es decir, las fibras musculares cardíacas están formadas por muchas células individuales
conectadas entre sí
en serie y en paralelo.
En
cada uno de los discos intercalados las membranas celulares se fusionan entre
sí para formar uniones «comunicantes»
(en hendidura) permeables que permiten una rápida difusión. Por tanto, desde un
punto de vista funcional los iones se mueven con facilidad en el líquido
intracelular a lo largo del eje longitudinal de las fibras musculares
cardíacas, de modo que los potenciales de acción viajan fácilmente desde una
célula muscular cardíaca a la siguiente, a través de los discos intercalados.
Por tanto, el músculo cardíaco es un sincitio
de muchas células musculares cardíacas en el que las células están tan interconectadas entre sí
que cuando una célula se excita el potencial de acción se propaga rápidamente a
todas.
El
corazón realmente está formado por dos sincitios: el sincitio
auricular, que forma las paredes de las
dos aurículas, y el sincitio ventricular,
que forma las paredes de los dos ventrículos. Las aurículas están separadas de
los ventrículos por tejido fibroso que rodea las aberturas de las válvulas auriculoventriculares (AV)entre
las aurículas y los ventrículos. Normalmente los potenciales no se conducen
desde el sincitio auricular hacia el sincitio ventricular directamente a través
de este tejido fibroso. Por el contrario, solo son conducidos por medio de un sistema de
conducción especializado denominado haz
AV, que es un fascículo de fibras de conducción de
varios milímetros de diámetro.
Esta división del músculo del corazón en dos sincitios funcionales permite que las aurículas se contraigan un pequeño intervalo antes de la contracción ventricular, lo que es importante para la eficacia del bombeo del corazón.
POTENCIALES
DE ACCIÓN EN EL MÚSCULO CARDÍACO
El potencial de acción que se registra en una fibra muscular ventricular es en promedio de aproximadamente 105 mV, lo que significa que el potencial intracelular aumenta desde un valor muy negativo, de aproximadamente –85 mV, entre los latidos hasta un valor ligeramente positivo, de aproximadamente +20 mV, durante cada latido. Después de la espiga inicial la membrana permanece despolarizada durante aproximadamente 0,2 s, mostrando una meseta, seguida al final de la meseta de una repolarización súbita. La presencia de esta meseta del potencial de acción hace que la contracción ventricular dure hasta 15 veces más en el músculo cardíaco que en el músculo esquelético.
¿QUÉ
PRODUCE EL POTENCIAL DE ACCIÓN PROLONGADO Y LA MESETA?
Dos
diferencias importantes entre las propiedades de la membrana del músculo
cardíaco y esquelético son responsables del potencial de acción prolongado y de
la meseta del músculo cardíaco.
Primero,
el potencial de acción del músculo esqueléticoestá
producido casi por completo por la apertura súbita de grandes números de
canales rápidos de sodio que permiten que grandes cantidades de iones sodio
entren en la fibra muscular esquelética desde el líquido extracelular. A estos
canales se les denomina canales «rápidos» porque permanecen abiertos solo
algunas milésimas de segundo y después se cierran súbitamente. Al final de este
cierre se produce la repolarización y el potencial de acción ha terminado en
otra milésima de segundo aproximadamente.
En
el músculo cardíaco, el potencial de acción está producido por la apertura de
dos tipos de canales: 1) los mismos canales rápidos de sodio
activados por el voltaje que en el músculo esquelético y
2) otra población totalmente distinta de canales de calcio de tipo L
(canales lentos de calcio), que también
se denominan canales de calcio-sodio.
Esta segunda población de canales difiere de los canales rápidos de sodio en
que se abren con mayor lentitud y, lo que es incluso más importante, permanecen
abiertos durante varias décimas de segundo. Durante este tiempo fluye una gran
cantidad de iones tanto calcio como sodio a través de estos canales hacia el
interior de la fibra muscular cardíaca, y esta actividad mantiene un período prolongado de
despolarización, dando lugar a la meseta del potencial de acción.
Además, los iones calcio que entran durante esta fase de meseta activan el
proceso contráctil del músculo, mientras que los iones calcio que producen la contracción
del músculo esquelético proceden del retículo sarcoplásmico intracelular.
La
segunda diferencia funcional importante entre el músculo cardíaco y el músculo
esquelético que ayuda a explicar tanto el potencial de acción prolongado como
su meseta es la siguiente: inmediatamente después del inicio del potencial de
acción, la permeabilidad
de la membrana del músculo cardíaco a los iones
potasio
disminuye aproximadamente cinco veces, un efecto
que no aparece en el músculo esquelético. Esta disminución de la permeabilidad
al potasio se puede deber al exceso de flujo de entrada de calcio
a través de los canales de calcio que se acaba de señalar.
Independientemente de la causa, la disminución de la permeabilidad al potasio reduce mucho el flujo de salida de iones potasio de carga positiva durante la meseta del potencial de acción y, por tanto, impide el regreso rápido del voltaje del potencial de acción a su nivel de reposo. Cuando los canales lentos de calcio-sodio se cierran después de 0,2 a 0,3 s y se interrumpe el flujo de entrada de iones calcio y sodio, también aumenta rápidamente la permeabilidad de la membrana a los iones potasio; esta rápida pérdida de potasio desde la fibra inmediatamente devuelve el potencial de membrana a su nivel de reposo, finalizando de esta manera el potencial de acción.
FASES DEL POTENCIAL DE ACCIÓN DEL MÚSCULO CARDÍACO
Fase 0 (despolarización),
los canales de sodio rápidos se abren. Cuando la célula cardíaca es estimulada
y se despolariza, el potencial de membrana se hace más positivo. Los canales de
sodio activados por el voltaje (canales de sodio rápidos)
se abren y permiten
que el sodio circule rápidamente hacia el interior de la célula y la
despolarice. El potencial de membrana
alcanza +20 mV aproximadamente
antes de que los canales de sodio se cierren.
Fase 1 (repolarización inicial),
los canales de sodio
rápidos se cierran. Los canales de sodio se cierran,
la célula empieza a repolarizarse y los iones potasiosalen de la célula a
través de los canales de potasio.
Fase 2 (meseta),
los canales de calcio se
abren y los canales de potasio rápidos se cierran.
Tiene lugar una breve repolarización inicial y el potencial de acción alcanza
una meseta como consecuencia de: 1) una mayor
permeabilidad a los iones calcio, y 2) la disminución de la permeabilidad a los iones potasio.
Los canales de calcio activados por el voltaje se abren lentamente durante las
fases 1 y 0, y el calcio entra en la célula. Después, los canales de potasio se
cierran, y la combinación de una reducción en la salida de iones potasio y un
aumento de la entrada de iones calcio lleva a que el potencial de acción
alcance una meseta.
Fase 3 (repolarización rápida),
los canales de calcio se
cierran y los canales de potasio lentos se abren.
El cierre de los canales iónicos de calcio y el aumento de la permeabilidad a
los iones potasio, que permiten que los iones potasio salgan rápidamente de la
célula, pone fin a la meseta
y devuelve el potencial de membrana de la célula a su nivel de reposo.
Fase 4 (potencial de membrana de reposo)
con valor medio aproximado de −90 mV.
VELOCIDAD
DE LA CONDUCCIÓN DE LAS SEÑALES EN EL MÚSCULO CARDÍACO
La velocidad de la conducción de la señal del potencial de acción excitador a lo largo de las fibras musculares auriculares y ventriculares es de unos 0,3 a 0,5 m/s, o aproximadamente 1/250 de la velocidad en las fibras nerviosas grandes y en torno a 1/10 de la velocidad en las fibras musculares esqueléticas. La velocidad de conducción en el sistema especializado de conducción del corazón, en las fibras de Purkinje, es de hasta 4 m/s en la mayoría de las partes del sistema, lo que permite una conducción razonablemente rápida de la señal excitadora hacia las diferentes partes del corazón.
PERÍODO
REFRACTARIO DEL MÚSCULO CARDÍACO
El
músculo cardíaco, al igual que todos los tejidos excitables, es refractario a
la reestimulación durante el potencial de acción. Por tanto, el período
refractario del corazón es el intervalo de tiempo durante el cual un
impulso cardíaco normal no puede reexcitar una zona ya excitada de músculo
cardíaco. El período refractario normal
del ventrículo es de 0,25 a 0,30 s,
que es aproximadamente la duración del potencial de acción en meseta prolongado.
Hay un período refractario relativo
adicional de aproximadamente 0,05 s,
durante el cual es
más difícil de lo normal excitar el músculo,
sin embargo, se
puede excitar con una señal excitadora muy intensa.
El
término «acoplamiento excitación-contracción»
se refiere al mecanismo mediante el cual el potencial de acción hace que las miofibrillas
del músculo se contraigan.
Al
igual que en el músculo esquelético, cuando un potencial de acción pasa sobre
la membrana del músculo cardíaco el potencial de acción se propaga hacia el
interior de la fibra muscular cardíaca a lo largo de las membranas de los túbulos
transversos (T). Los potenciales de acción de
los túbulos T, a su vez, actúan sobre las membranas de los túbulos
sarcoplásmicos longitudinales para producir la liberación de iones calcio hacia
el sarcoplasma muscular desde el retículo sarcoplásmico.
En algunas milésimas de segundo más estos iones calcio difunden hacia las
miofibrillas y catalizan las reacciones químicas que favorecen el deslizamiento
de los filamentos de actina y de miosina entre sí, lo que da lugar a la
contracción muscular.
Hasta
ahora este mecanismo de acoplamiento excitación-contracción es el mismo que el
del músculo esquelético, aunque hay un segundo efecto que es bastante
diferente. Además de los iones calcio que se liberan hacia el sarcoplasma desde
las cisternas del retículo sarcoplásmico, también difunde una gran cantidad de iones calcio
adicionales hacia el sarcoplasma desde los propios túbulos T en el momento
del potencial de acción, que abre los canales de calcio dependientes del voltaje
a la membrana del túbulo T.
El
calcio que entra en la célula activa después los canales
de liberación de calcio, también denominados canales de receptor de rianodina,
en la membrana del retículo sarcoplásmico, para activar la liberación de calcio
en el sarcoplasma. Los iones calcio en el sarcoplasma interaccionan después con
la troponina para iniciar la formación y contracción de puente transversal
mediante el mismo mecanismo básico que se ha descrito para el músculo
esquelético.
Sin
el calcio procedente de los túbulos T la fuerza de la contracción del músculo
cardíaco se reduciría de manera considerable porque el retículo
sarcoplásmico
del músculo cardíaco está peor desarrollado
que el del músculo esquelético y no almacena suficiente calcio para generar una
contracción completa. No obstante, los túbulos
T del músculo cardíaco tienen un diámetro cinco veces mayor que los túbulos
del músculo esquelético, lo que significa un volumen 25
veces mayor. Además, en el interior de los túbulos T hay una gran cantidad de mucopolisacáridos que tienen carga negativa y que se unen a una abundante
reserva de iones calcio, manteniéndolos disponibles para
su difusión hacia el interior de la fibra muscular cardíaca cuando aparece un
potencial de acción en un túbulo T.
La
fuerza de la contracción del músculo cardíaco depende en gran medida de la concentración de iones calcio en los líquidos
extracelulares.
Al final de la meseta del potencial de acción cardíaco se interrumpe súbitamente el flujo de entrada de iones calcio hacia el interior de la fibra muscular y los iones calcio del sarcoplasma se bombean rápidamente hacia el exterior de las fibras musculares, hacia el retículo sarcoplásmico y hacia el espacio de los túbulos T-líquido extracelular. El transporte de calcio de nuevo al retículo sarcoplásmico se consigue con la ayuda de una bomba de calcio adenosina trifosfatasa (ATPasa).
DURACIÓN DE LA CONTRACCIÓN
El
músculo cardíaco comienza a contraerse algunos milisegundos después de la
llegada del potencial de acción y sigue contrayéndose hasta algunos
milisegundos después de que finalice. Por tanto, la duración de la contracción
del músculo cardíaco depende principalmente de la duración del potencial de
acción, incluyendo la meseta,
aproximadamente 0,2 s en el músculo
auricular y 0,3 s en el músculo
ventricular.
Los
fenómenos cardíacos que se producen desde el comienzo de un latido cardíaco
hasta el comienzo del siguiente se denominan ciclo
cardíaco. Cada ciclo es iniciado por la generación
espontánea de un potencial de acción en el nódulo
sinusal.
Este nódulo está localizado en la pared superolateral de la aurícula derecha, cerca del orificio de la vena cava superior, y el potencial de acción viaja desde aquí rápidamente por ambas aurículas y después a través del haz AV hacia los ventrículos. Debido a esta disposición especial del sistema de conducción desde las aurículas hacia los ventrículos, hay un retraso de más de 0,1 s durante el paso del impulso cardíaco desde las aurículas a los ventrículos. Esto permite que las aurículas se contraigan antes de la contracción ventricular, bombeando de esta manera sangre hacia los ventrículos antes de que comience la intensa contracción ventricular. Por tanto, las aurículas actúan como bombas de cebado para los ventrículos, y los ventrículos a su vez proporcionan la principal fuente de potencia para mover la sangre a través del sistema vascular del cuerpo.
DIÁSTOLE
Y SÍSTOLE
El ciclo cardíaco está formado por un período de relajación que se denomina diástole, seguido de un período de contracción denominado sístole.
La
duración del ciclo cardíaco total, incluidas la sístole y la diástole, es el valor inverso de la frecuencia cardíaca.
Por ejemplo, si la frecuencia cardíaca es de 72 latidos/min, la duración del
ciclo cardíaco es de 1/72 min/latido, aproximadamente 0,0139 min por latido, o
0,833 s por latido.
EL AUMENTO DE LA FRECUENCIA CARDÍACA REDUCE LA DURACIÓN DEL CICLO CARDÍACO
Cuando
aumenta la frecuencia cardíaca, la duración de cada ciclo cardíaco disminuye, incluidas las
fases de contracción y relajación. La
duración del potencial de acción y el período de contracción (sístole) también
decrece, aunque no en un porcentaje tan elevado como en la fase de relajación (diástole).
Para una frecuencia cardíaca normal de 72 latidos/min, la sístole comprende aproximadamente
0,4 del ciclo cardíaco completo. Para una frecuencia cardíaca triple de lo
normal, la sístole supone aproximadamente 0,65 del ciclo cardíaco completo. Esto
significa que el corazón que late a una frecuencia muy rápida no permanece
relajado el tiempo suficiente para permitir un llenado completo de las cámaras
cardíacas antes de la siguiente contracción.
RELACIÓN DEL ELECTROCARDIOGRAMA CON EL CICLO CARDÍACO
El
electrocardiograma muestra las ondas P, Q, R, S y T. Son los voltajes
eléctricos que genera el corazón, y son registrados mediante el electrocardiógrafo
desde la superficie del cuerpo.
La
onda P está
producida por la propagación de la despolarización
en las aurículas, y es seguida por la
contracción auricular, que produce una ligera elevación de la curva
de presión auricular inmediatamente
después de la onda P electrocardiográfica.
Aproximadamente
0,16 s después del inicio de la onda P, las ondas
QRS aparecen como consecuencia de la despolarización eléctrica de los ventrículos,
que inicia la contracción de los ventrículos y hace que comience a elevarse
la presión ventricular. Por tanto,
el complejo QRScomienza un poco antes del inicio de la sístole
ventricular.
Finalmente,
la onda T ventricular
representa la fase de repolarización
de los ventrículos, cuando las fibras del músculo
ventricular comienzan a relajarse. Por tanto, la onda T se produce un poco
antes del final de la contracción ventricular.
FUNCIÓN
DE LAS AURÍCULAS COMO BOMBAS DE CEBADO PARA LOS VENTRÍCULOS
La
sangre normalmente fluye de forma continua desde las grandes venas hacia las
aurículas; aproximadamente el 80%
de la sangre fluye directamente a través de las aurículas
hacia los ventrículos
incluso antes de que se contraigan las aurículas. Después, la contracción
auricular habitualmente produce un llenado de un 20%
adicional de los ventrículos. Por tanto, las aurículas actúan como bombas de
cebado que aumentan la eficacia del bombeo ventricular hasta un 20%. Sin embargo,
el corazón puede seguir funcionando en la mayor parte de las condiciones incluso
sin esta eficacia de un 20% adicional porque normalmente tiene la capacidad de
bombear entre el 300 y el 400% más de sangre de la que necesita el cuerpo en
reposo.
CAMBIOS
DE PRESIÓN EN LAS AURÍCULAS: LAS ONDAS A, C Y V
En la curva de presión auricular
se muestran tres pequeñas elevaciones de presión, denominadas curvas de presión
auricular a, c y v.
La onda a está producida por la contracción auricular. Habitualmente la presión auricular
derecha aumenta de 4 a 6 mmHg durante la contracción auricular y la presión
auricular izquierda aumenta aproximadamente de 7 a 8 mmHg.
La onda c se produce cuando los ventrículos comienzan
a contraerse; está producida en parte por un ligero flujo retrógrado de sangre
hacia las aurículas al comienzo de la contracción ventricular, pero
principalmente por la protrusión de las válvulas AV retrógradamente hacia las
aurículas debido al aumento de presión de los ventrículos.
La onda v se produce hacia el final de la
contracción ventricular; se debe al flujo lento de sangre hacia las aurículas desde las venas
mientras las válvulas AV están cerradas durante la contracción ventricular.
Después, cuando ya ha finalizado la contracción ventricular, las válvulas AV se
abren, y permiten que esta sangre auricular almacenada fluya rápidamente hacia
los ventrículos, lo que hace que la onda v desaparezca.
LOS VENTRÍCULOS SE LLENAN DE
SANGRE DURANTE LA DIÁSTOLE
Durante la sístole ventricular se
acumulan grandes cantidades de sangre en las aurículas derecha e izquierda
porque las válvulas AV están cerradas. Por tanto, tan pronto como ha finalizado
la sístole y las presiones ventriculares disminuyen de nuevo a sus valores
diastólicos bajos, el aumento moderado de presión que se ha generado en las
aurículas durante la sístole ventricular inmediatamente abre las válvulas AV y
permite que la sangre fluya rápidamente hacia los ventrículos, como se muestra
en la elevación de la curva de volumen ventricular izquierdo. Es el denominado período dellenado rápido de los ventrículos.
El período de llenado rápido dura aproximadamente
el primer tercio de la diástole. Durante el tercio medio de la diástole
normalmente solo fluye una pequeña cantidad de sangre hacia los ventrículos; esta
es la sangre que continúa drenando hacia las aurículas desde las venas y que
pasa a través de las aurículas directamente hacia los ventrículos.
Durante el último tercio de la diástole las
aurículas se contraen y aportan un impulso adicional al flujo de entrada de sangre
hacia los ventrículos. Este mecanismo es responsable de aproximadamente el 20%
del llenado de los ventrículos durante cada ciclo cardíaco.
PERÍODO
DE CONTRACCIÓN ISOVOLUMÉTRICA (ISOMÉTRICA)
Inmediatamente después del comienzo de la
contracción ventricular se produce un aumento súbito de presión ventricular, lo
que hace que se cierren las válvulas AV.
Después son necesarios otros 0,02 a 0,03 s para que el ventrículo acumule una presión suficiente para abrir las válvulas AV semilunares (aórtica y pulmonar) contra las presiones de la aorta y de la arteria pulmonar. Por tanto, durante este período se produce contracción en los ventrículos, pero no se produce vaciado. Es el llamado período de contracción isovolumétrica o isométrica, lo que quiere decir que se produce aumento de la tensión en el músculo cardíaco, pero con un acortamiento escaso o nulo de las fibras musculares.
PERÍODO DE EYECCIÓN
Cuando la presión
ventricular izquierda aumenta ligeramente por encima de 80 mmHg (y la presión ventricular
derecha ligeramente por encima de 8 mmHg), las presiones ventriculares abren
las válvulas semilunares. Inmediatamente comienza a salir la sangre de los
ventrículos.
Aproximadamente el 60% de
la sangre del ventrículo al final de la diástole es expulsada durante la sístole;
en torno al 70% de esta porción es expulsado durante el primer tercio del
período de eyección y el 30% restante del vaciado se produce durante los dos
tercios siguientes. Por tanto, el primer tercio se denomina período de eyección
rápida y los dos tercios finales período de eyección lenta.
PERÍODO DE RELAJACIÓN
ISOVOLUMÉTRICA (ISOMÉTRICA)
Al final de la sístole comienza súbitamente la relajación ventricular, lo que permite que las presiones intraventriculares derecha e izquierda disminuyan rápidamente. Las presiones elevadas de las grandes arterias distendidas que se acaban de llenar con la sangre que procede de los ventrículos que se han contraído empujan inmediatamente la sangre de nuevo hacia los ventrículos, lo que cierra súbitamente las válvulas aórtica y pulmonar. Durante otros 0,03 a 0,06 s el músculo cardíaco sigue relajándose, aun cuando no se modifica el volumen ventricular, dando lugar al período de relajación isovolumétrica o isométrica. Durante este período las presiones intraventriculares disminuyen rápidamente y regresan a sus bajos valores diastólicos. Después se abren las válvulas AV para comenzar un nuevo ciclo de bombeo ventricular.
VOLUMEN TELEDIASTÓLICO,
VOLUMEN TELESISTÓLICO Y VOLUMEN SISTÓLICO
Durante la diástole, el
llenado normal de los ventrículos aumenta el volumen de cada uno de los ventrículos
hasta aproximadamente 110 a 120 ml. Este volumen se denomina volumen
telediastólico.
Después, a medida que los
ventrículos se vacían durante la sístole, el volumen disminuye aproximadamente
70 ml, lo que se denomina volumen sistólico. El volumen restante que queda en cada
uno de los ventrículos, aproximadamente 40 a 50 ml, se denomina volumen
telesistólico. La fracción del volumen telediastólico que es propulsada se denomina
fracción de eyección, que habitualmente es igual a 0,6 (o el 60%)
aproximadamente.
Cuando el corazón se
contrae con fuerza el volumen telesistólico puede disminuir hasta un valor tan bajo
como 10 a 20 ml. Por el contrario, cuando fluyen grandes cantidades de sangre
hacia los ventrículos durante la diástole, los volúmenes telediastólicos ventriculares
pueden llegar a ser tan grandes como 150 a 180 ml en el corazón sano. Mediante
el aumento del volumen telediastólico y la reducción del volumen telesistólico
se puede aumentar el volumen sistólico hasta más del doble de lo normal.
VÁLVULAS
AURICULOVENTRICULARES
Las válvulas AV (las válvulas tricúspide y mitral) impiden el flujo retrógrado de sangre desde los ventrículos hacia las aurículas durante la sístole, y las válvulas semilunares (es decir, las válvulas aórtica y de la arteria pulmonar) impiden el flujo retrógrado desde las arterias aorta y pulmonar hacia los ventrículos durante la diástole. Estas válvulas, para el ventrículo izquierdo, se cierran y abren pasivamente. Es decir, se cierran cuando un gradiente de presión retrógrada empuja la sangre hacia atrás, y se abren cuando un gradiente de presión anterógrada fuerza la sangre en dirección anterógrada. Por motivos anatómicos, las válvulas AV, que están formadas por una película delgada, casi no precisan ningún flujo retrógrado para cerrarse, mientras que las válvulas semilunares, que son mucho más fuertes, precisan un flujo retrógrado bastante rápido durante algunos milisegundos.
FUNCIÓN DE LOS MÚSCULOS PAPILARES
Los músculos papilares que se unen a los velos de las válvulas AV mediante
las cuerdas
tendinosas. Los músculos papilares se contraen cuando se contraen
las paredes ventriculares, pero, al contrario de lo que se podría esperar, no
contribuyen al cierre de las válvulas.
Por el contrario, tiran de los velos de las
válvulas hacia dentro, hacia los ventrículos, para impedir que protruyan
demasiado hacia las aurículas durante la contracción ventricular.
VÁLVULAS
AÓRTICA Y DE LA ARTERIA PULMONAR
Las válvulas semilunares aórtica y pulmonar
funcionan de una manera bastante distinta de las válvulas AV. Primero, las elevadas
presiones de las arterias al final de la sístole hacen que las válvulas semilunares
se cierren súbitamente, a diferencia del cierre mucho más suave de las
válvulas AV.
Segundo, debido a sus orificios más pequeños, la
velocidad de la eyección de la sangre a través de las válvulas aórtica y
pulmonar es mucho mayor que a través de las válvulas AV, que son mucho mayores.
Además, debido al cierre rápido y a la eyección rápida, los bordes de las
válvulas aórtica y pulmonar están sometidos a una abrasión mecánica mucho mayor
que las válvulas AV. Las válvulas AV tienen el soporte de las cuerdas
tendinosas, lo que no ocurre en el caso de las válvulas semilunares. A partir
de la anatomía de las válvulas aórtica y pulmonar es evidente que deben estar situadas
sobre una base de un tejido fibroso especialmente fuerte, pero muy flexible
para soportar las tensiones físicas adicionales.
Cuando el ventrículo izquierdo se contrae, la presión ventricular aumenta rápidamente hasta que se abre la válvula aórtica. Posteriormente, después de que se haya abierto la válvula, la presión del ventrículo aumenta mucho menos rápidamente, porque la sangre sale inmediatamente del ventrículo hacia la aorta y después hacia las arterias de distribución sistémica.
La entrada de sangre en las arterias durante la
sístole hace que sus paredes se distiendan y que la presión aumente hasta
aproximadamente 120 mmHg.
Al final de la sístole, después de que el
ventrículo izquierdo haya dejado de impulsar sangre y se haya cerrado la
válvula aórtica, las paredes elásticas de las arterias mantienen una presión
elevada en las arterias, incluso durante la diástole.
Se produce una incisura en la curva de presión
aórtica cuando se cierra la válvula aórtica; está producida por un corto
período de flujo retrógrado de sangre inmediatamente antes del cierre de la válvula,
seguido por la interrupción súbita del flujo retrógrado.
Después de que se haya cerrado la válvula aórtica,
la presión en el interior de la aorta disminuye lentamente durante toda la
sístole porque la sangre que está almacenada en las arterias elásticas distendidas
fluye continuamente a través de los vasos periféricos de nuevo hacia las venas.
Antes de que se contraiga de nuevo el ventrículo, la presión aórtica habitualmente
ha disminuido hasta aproximadamente 80
mmHg (presión diastólica), que es dos tercios de la presión máxima de 120 mmHg (presión sistólica) que
se produce en la aorta durante la contracción ventricular.
RELACIÓN
DE LOS TONOS CARDÍACOS CON EL BOMBEO CARDÍACO
Cuando se ausculta el corazón con un estetoscopio
no se oye la apertura de las válvulas porque este es un proceso relativamente
lento que no suele hacer ruido. Sin embargo, cuando las válvulas se cierran, los
velos de las válvulas y los líquidos circundantes vibran bajo la influencia de
los cambios súbitos de presión, generando un sonido que viaja en todas las
direcciones a través del tórax.
Cuando se contraen
los ventrículos primero se oye un ruido que está producido por el cierre de las
válvulas AV. El tono de la vibración es bajo y relativamente prolongado,
y se conoce como el primer tono cardíaco. Cuando se cierran las
válvulas aórtica y pulmonar al final de la sístole se oye un golpe seco
y rápido porque estas válvulas se cierran rápidamente, y los líquidos
circundantes vibran durante un período corto. Este sonido se denomina segundo tono
cardíaco.
GENERACIÓN DE TRABAJO DEL CORAZÓN
El trabajo sistólico del corazón es la cantidad de energía que el corazón convierte en trabajo durante cada latido cardíaco mientras bombea sangre hacia las arterias. El trabajo minuto es la cantidad total de energía que se convierte en trabajo en 1 min; este parámetro es igual al trabajo sistólico multiplicado por la frecuencia cardíaca por minuto.
El trabajo del corazón se utiliza de dos maneras.
Primero, la mayor proporción se utiliza, con mucho, para mover la sangre desde
las venas de baja presión hacia las arterias de alta presión. Esto se denomina
trabajo volumen-presión o trabajo externo. Segundo, una pequeña proporción de
la energía se utiliza para acelerar la sangre hasta su velocidad de eyección a
través de las válvulas aórtica y pulmonar, que es el componente de energía
cinética del flujo sanguíneo del trabajo cardíaco.
El trabajo externo del ventrículo derecho es
normalmente de alrededor de la sexta parte del trabajo del ventrículo izquierdo
debido a la diferencia de seis veces de las presiones sistólicas que bombean los
dos ventrículos. El trabajo adicional de cada uno de los ventrículos necesario
para generar la energía cinética del flujo sanguíneo es proporcional a la masa
de sangre que se expulsa multiplicada por el cuadrado de la velocidad de
eyección.
La curva
de presión diastólica se determina llenando el corazón con volúmenes de
sangre progresivamente mayores y midiendo la presión diastólica inmediatamente
antes de que se produzca la contracción ventricular, que es la presión telediastólica del ventrículo.
La curva de presión sistólica se determina
registrando la presión sistólica que se alcanza durante la contracción
ventricular a cada volumen de llenado.
Hasta que el volumen del ventrículo que no se está
contrayendo no aumenta por encima de aproximadamente 150 ml, la presión «diastólica» no aumenta
mucho. Por tanto, hasta este volumen la sangre puede fluir con facilidad hacia
el ventrículo desde la aurícula. Por encima de 150 ml la presión diastólica
ventricular aumenta rápidamente, en parte porque el tejido fibroso del corazón
ya no se puede distender más y en parte porque el pericardio que rodea el corazón
se ha llenado casi hasta su límite.
Durante la contracción ventricular, la presión «sistólica» aumenta
incluso a volúmenes ventriculares bajos y alcanza un máximo a un volumen
ventricular de 150 a 170 ml. Después, a medida que sigue aumentando el
volumen, la presión sistólica llega a disminuir en algunas situaciones, como se
muestra por la disminución de la curva de presión sistólica, porque a estos
volúmenes elevados los filamentos de actina y de miosina de las fibras
musculares cardíacas están tan separados que la fuerza de la contracción de
cada una de las fibras cardíacas se hace menos óptima.
La presión sistólica máxima del ventrículo izquierdo normal está entre 250 y 300 mmHg, aunque esto varía mucho con la fuerza del corazón de cada persona y con el grado de la estimulación del corazón por los nervios cardíacos. Para el ventrículo derecho normal la presión sistólica máxima está entre 60 y 80 mmHg.
«DIAGRAMA
VOLUMEN-PRESIÓN» DURANTE EL CICLO CARDÍACO: TRABAJO CARDÍACO
El diagrama
volumen-presión forman un bucle del ciclo cardíaco para la función
normal del ventrículo izquierdo. Está dividido en cuatro fases.
Fase I: período
de llenado. La fase I del diagrama volumen-presión comienza a un
volumen ventricular de aproximadamente 50 ml y una presión diastólica próxima a
2 a 3 mmHg. La cantidad de sangre que queda en el ventrículo después del latido
previo, 50 ml, se denomina volumen telesistólico.
A medida que la sangre venosa fluye hacia el
ventrículo desde la aurícula izquierda, el volumen ventricular normalmente
aumenta hasta aproximadamente 120 ml, el denominado volumen telediastólico, un
aumento de 70 ml.
Fase II: período
de contracción isovolumétrica. Durante la contracción isovolumétrica el
volumen del ventrículo no se modifica porque todas las válvulas están cerradas.
Sin embargo, la presión en el interior del ventrículo aumenta hasta igualarse a
la presión que hay en la aorta, hasta un valor de presión de aproximadamente 80
mmHg.
Fase III: período
de eyección. Durante la eyección la presión sistólica aumenta
incluso más debido a una contracción aún más intensa del ventrículo. Al mismo
tiempo, el volumen del ventrículo disminuye porque la válvula aórtica ya se ha
abierto y la sangre sale del ventrículo hacia la aorta.
Fase IV: período
de relajación isovolumétrica. Al final del período de eyección se
cierra la válvula aórtica, y la presión ventricular disminuye
de nuevo hasta el nivel de la presión diastólica.
El área que encierra este diagrama volumen-presión
funcional (la zona sombreada denominada TE) representa el trabajo cardíaco externo
neto del ventrículo durante su ciclo de contracción. En estudios experimentales
de contracción cardíaca este diagrama se utiliza para calcular el trabajo
cardíaco.
Cuando el corazón bombea grandes cantidades de
sangre, el área del diagrama de trabajo se hace mucho mayor. Es decir, se
extiende más hacia la derecha porque el ventrículo se llena con más sangre durante
la diástole, se eleva mucho más porque el ventrículo se contrae con mayor
presión, y habitualmente se extiende más a la izquierda porque el ventrículo se
contrae hasta un menor volumen, especialmente si el sistema nervioso simpático
estimula un aumento de actividad del ventrículo.
CONCEPTOS
DE PRECARGA Y POSCARGA
Cuando se evalúan las propiedades contráctiles del
músculo es importante especificar el grado de tensión del músculo cuando
comienza a contraerse, que se denomina precarga,
y especificar la carga contra la que el músculo ejerce su fuerza contráctil,
que se denomina poscarga.
Para la contracción cardíaca habitualmente se
considera que la precarga es la presión telediastólica cuando el ventrículo ya se
ha llenado.
La poscarga del ventrículo es la presión de la
aorta que sale del ventrículo.
La importancia de los conceptos de precarga y poscarga es que en muchos estados funcionales anormales del corazón o de la circulación, la presión durante el llenado del ventrículo (la precarga), la presión arterial contra la que se debe contraer el ventrículo (la poscarga) o ambas están alteradas con respecto a su situación normal en un grado importante.
ENERGÍA
QUÍMICA NECESARIA PARA LA CONTRACCIÓN CARDÍACA: LA UTILIZACIÓN DE OXÍGENO POR
EL CORAZÓN
El músculo cardíaco, al igual que el músculo esquelético, utiliza energía química para realizar el trabajo de la contracción. Aproximadamente el 70-90% de esta energía procede normalmente del metabolismo oxidativo de los ácidos grasos, donde el 10-30%, aproximadamente, procede de otros nutrientes, especialmente lactato y glucosa. Por tanto, la velocidad del consumo de oxígeno por el miocardio es una medida excelente de la energía química que se libera mientras el corazón realiza su trabajo.
El consumo de oxígeno ha demostrado ser también
casi proporcional a la tensión que se produce en el músculo cardíaco durante la
contracción multiplicada por la duración de tiempo durante la cual persiste la
contracción, denominada índice de tensión-tiempo. Como la tensión es alta
cuando lo es la presión sistólica, en correspondencia se usa más oxígeno. Además,
se gasta mucha más energía química a presiones sistólicas normales cuando el
ventrículo está dilatado anómalamente debido a que la tensión del músculo
cardíaco durante la contracción es proporcional a la presión multiplicada por
el diámetro del ventrículo. Esto se hace especialmente importante en caso de
insuficiencia cardíaca en la que el ventrículo cardíaco está dilatado y,
paradójicamente, la cantidad de energía química necesaria para una cantidad
dada de trabajo cardíaco es mayor de lo normal incluso cuando el corazón ya
está desfalleciendo.
EFICIENCIA
DE LA CONTRACCIÓN CARDÍACA
Durante la contracción del músculo cardíaco la
mayor parte de la energía química que se gasta se convierte en calor y
una porción mucho menor en trabajo. El cociente del trabajo
respecto al gasto de energía química total se denomina eficiencia de la contracción cardíaca, o simplemente eficiencia del corazón. La eficiencia
máxima del corazón normal está entre el 20
y el 25%. En personas con insuficiencia cardíaca este valor puede
disminuir hasta el 5-10%.
Cuando una persona está en reposo el corazón solo
bombea de 4 a 6 l de sangre
cada minuto. Durante el ejercicio intenso puede ser necesario que el corazón
bombee de cuatro a siete veces esta cantidad.
Los mecanismos básicos mediante los que se regula el volumen que bombea el corazón son: 1) regulación cardíaca intrínseca del bombeo en respuesta a los cambios del volumen de la sangre que fluye hacia el corazón, y 2) control de la frecuencia cardíaca y del bombeo cardíaco por el sistema nervioso autónomo.
REGULACIÓN
INTRÍNSECA DEL BOMBEO CARDÍACO: EL MECANISMO DE FRANK-STARLING
La cantidad de sangre que bombea el corazón cada minuto está determinada en gran medida, por lo común, por la velocidad del flujo sanguíneo hacia el corazón desde las venas, que se denomina retorno venoso. Es decir, todos los tejidos periféricos del cuerpo controlan su propio flujo sanguíneo local, y todos los flujos tisulares locales se combinan y regresan a través de las venas hacia la aurícula derecha. El corazón, a su vez, bombea automáticamente hacia las arterias esta sangre que le llega, de modo que pueda fluir de nuevo por el circuito.
Esta capacidad intrínseca del corazón de adaptarse
a volúmenes crecientes de flujo sanguíneo de entrada se denomina mecanismo de Frank-Starling del
corazón en honor de Otto Frank y Ernest Starling, dos grandes fisiólogos de
hace un siglo. Básicamente, el mecanismo de Frank-Starling significa que cuanto
más se distiende el músculo cardíaco durante el llenado, mayor es la fuerza de contracción
y mayor es la cantidad de sangre que bombea hacia la aorta. O, enunciado de
otra manera, dentro de límites fisiológicos el corazón bombea toda la
sangre que le llega procedente de las venas.
¿CUÁL
ES LA EXPLICACIÓN DEL MECANISMO DE FRANK-STARLING?
Cuando una cantidad adicional de sangre fluye hacia
los ventrículos, el propio músculo cardíaco es distendido hasta una mayor
longitud. Esta distensión, a su vez, hace que el músculo se contraiga con más
fuerza porque los filamentos de actina y de miosina son desplazados hacia un
grado más óptimo de superposición para la generación de fuerza. Por tanto, el
ventrículo, debido al aumento de la función de bomba, bombea automáticamente la
sangre adicional hacia las arterias.
Esta capacidad del músculo distendido, hasta una longitud óptima, de contraerse con un aumento del trabajo cardíaco, es característica de todo el músculo estriado. Además del importante efecto del aumento de longitud del músculo cardíaco, hay otro factor que aumenta la función de bomba del corazón cuando aumenta su volumen. La distensión de la pared de la aurícula derecha aumenta directamente la frecuencia cardíaca en un 10-20%, lo que también contribuye a aumentar la cantidad de sangre que se bombea cada minuto, aunque su contribución es mucho menor que la del mecanismo de Frank-Starling.
CURVAS
DE FUNCIÓN VENTRICULAR
Una de las mejores formas de expresar la capacidad
funcional de los ventrículos de bombear sangre es mediante las curvas de función ventricular.
Un tipo de curva de función ventricular denominada curva de
trabajo sistólico. Obsérvese que a medida que aumenta la presión auricular
de cada uno de los lados del corazón, el trabajo sistólico de ese lado aumenta
hasta que alcanza el límite de la capacidad de bombeo del ventrículo.
Otro tipo de curva de función ventricular
denominada curva de volumen ventricular. Las dos curvas representan la
función de los ventrículos del corazón humano basadas en datos extrapolados de
estudios con animales experimentales. A medida que aumentan las presiones las
aurículas derecha e izquierda, también lo hacen los volúmenes ventriculares por
minuto respectivos.
Así, las curvas de función ventricular son otra
forma de expresar el mecanismo de Frank-Starling del corazón. Es decir, a
medida que los ventrículos se llenan en respuesta a unas presiones auriculares más
altas, se produce aumento del volumen de los dos ventrículos y de la fuerza de
la contracción del músculo cardíaco, lo que hace que el corazón bombee mayores
cantidades de sangre hacia las arterias.
CONTROL
DEL CORAZÓN POR LOS NERVIOS SIMPÁTICOS Y PARASIMPÁTICOS
La eficacia de la función de bomba del corazón también está controlada por los nervios simpáticos y parasimpáticos (vagos), que inervan de forma abundante el corazón.
Para niveles dados de presión auricular de entrada, la cantidad de sangre que se bombea cada minuto (gasto cardíaco) con frecuencia se puede aumentar más de un 100% por la estimulación simpática. Por el contrario, el gasto se puede disminuir hasta un valor tan bajo como cero o casi cero por la estimulación vagal (parasimpática).
MECANISMOS
DE EXCITACIÓN DEL CORAZÓN POR LOS NERVIOS SIMPÁTICOS
La estimulación simpática intensa puede aumentar la
frecuencia cardíaca en seres humanos adultos jóvenes desde la frecuencia normal
de 70 latidos/min hasta 180 a 200 y, raras veces, incluso 250 latidos/min.
Además, la estimulación simpática aumenta la fuerza de la contracción cardíaca hasta
el doble de lo normal, aumentando de esta manera el volumen de sangre que se
bombea y aumentando la presión de eyección. Así, con frecuencia la estimulación
simpática puede aumentar el gasto cardíaco máximo hasta dos o tres veces,
además del aumento del gasto que produce el mecanismo de Frank-Starling que ya
se ha comentado.
Por el contrario, la inhibición de los nervios simpáticos
del corazón puede disminuir la función de bomba del corazón en un grado
moderado: en condiciones normales, las fibras nerviosas simpáticas que llegan
al corazón descargan continuamente a una frecuencia baja que mantiene el bombeo
aproximadamente un 30% por encima del que habría sin estimulación simpática.
LA ESTIMULACIÓN PARASIMPÁTICA (VAGAL) REDUCE LA FRECUENCIA CARDÍACA Y LA FUERZA DE LA CONTRACCIÓN
La estimulación intensa de las fibras nerviosas parasimpáticas de los nervios vagos que llegan al corazón puede interrumpir el latido cardíaco durante algunos segundos, pero después el corazón habitualmente «escapa» y late a una frecuencia de 20 a 40 latidos/min mientras continúe la estimulación parasimpática. Además, la estimulación vagal intensa puede reducir la fuerza de la contracción del músculo cardíaco en un 20-30%.
Las fibras
vagales se distribuyen principalmente por las aurículas y no
mucho en los ventrículos, en los que se produce la contracción de potencia del
corazón. Esta distribución explica por qué el efecto de la estimulación vagal
tiene lugar principalmente sobre la reducción de la frecuencia cardíaca, en lugar
de reducir mucho la fuerza de la contracción del corazón. Sin embargo, la gran
disminución de la frecuencia cardíaca, combinada con una ligera reducción de la
fuerza de la contracción cardíaca, puede reducir el bombeo ventricular en un
50% o más.
EFECTO
DE LA ESTIMULACIÓN SIMPÁTICA Y PARASIMPÁTICA SOBRE LA CURVA DE FUNCIÓN CARDÍACA
A cualquier presión auricular derecha dada, el gasto cardíaco aumenta durante el aumento de la estimulación simpática y disminuye durante el aumento de la estimulación parasimpática. Estas modificaciones del gasto que se producen por la estimulación del sistema nervioso autónomo se deben tanto a modificaciones de la frecuencia cardíaca como a modificaciones de la fuerza contráctil del corazón.
EFECTO
DE LOS IONES POTASIO
El exceso
de potasio hace que el corazón esté dilatado y flácido, y también
reduce la frecuencia cardíaca. Grandes cantidades de potasio también
pueden bloquear la conducción del impulso cardíaco desde las aurículas hacia
los ventrículos a través del haz AV.
La elevación de la concentración de potasio hasta solo 8 a 12 mEq/l (dos a tres
veces el valor normal) puede producir una profunda debilidad del corazón, una
alteración del ritmo e incluso la muerte.
Estos efectos se deben parcialmente al hecho de que
una concentración elevada de potasio en los líquidos extracelulares reduce el
potencial de membrana en reposo de las fibras del músculo cardíaco.
EFECTO
DE LOS IONES CALCIO
Un exceso
de iones calcio produce efectos casi exactamente contrarios a los de
los iones potasio, haciendo que el corazón progrese hacia una contracción
espástica. Este efecto está producido por el efecto directo de los
iones calcio en el inicio del proceso contráctil cardíaco.
El aumento de la temperatura corporal, como ocurre durante
la fiebre, produce un gran aumento de la frecuencia cardíaca, a veces hasta del
doble del valor normal. El descenso de la temperatura produce una gran
disminución de la frecuencia cardíaca, que puede disminuir hasta solo algunos
latidos por minuto cuando una persona está cerca de la muerte por hipotermia en
el intervalo de temperatura corporal de 16 °C a 21 °C. Estos efectos
probablemente se deben al hecho de que el calor aumenta la permeabilidad de la
membrana del músculo cardíaco a los iones que controlan la frecuencia cardíaca,
acelerando el proceso de autoexcitación.
La fuerza contráctil del corazón con frecuencia se incrementa transitoriamente cuando hay un aumento moderado de la temperatura, como ocurre durante el ejercicio corporal, aunque una elevación prolongada de la temperatura agota los sistemas metabólicos del corazón y finalmente produce debilidad.
El aumento de la presión arterial
en la aorta no reduce el gasto cardíaco hasta que la presión arterial media
aumenta por encima de aproximadamente 160 mmHg. En otras palabras, durante la
función normal del corazón a presiones arteriales sistólicas normales (80 a 140
mmHg) el gasto cardíacoestá determinado casi totalmente por la facilidad
del flujo sanguíneo a través de los tejidos corporales, que a su vez controla
el retorno
venoso de la sangre hacia el corazón.
La unión neuromuscular es la unión entre una de las fibras nerviosas de neuronas motoras grandes provenientes de las astas anteriores de la médula espinal que pasan por el vientre muscular y una de las fibras del músculo esquelético (solo el 2% de las fibras musculares omiten esta regla).
Esta fibra nerviosa se une al punto medio de la fibra muscular y forma un complejo de terminaciones nerviosas que se acoplan con la fibra muscular, esta estructura se llama placa motora terminal, que está envuelta por células de Schwann que aísla la placa de los líquidos extracelulares.
Cuando se genera una potencia de acción, este inicia en el centro y se extiende a los extremos de la fibra muscular. La membrana que ha sido invaginada se denomina gotiera sináptica o valle sináptico, en este hay pliegues pequeños de la membrada de la fibra muscular llamadas hendiduras subneurales, su función es aumentar el área en la que el potencial de acción puede actuar. El espacio que existe entre la vesícula sináptica y la gotiera se llama espacio sináptico o hendidura sináptica y mide de 20 a 30 nm de ancho. El ATP es la fuente de energía que se usa para producir acetilcolina, que es el transmisor que excita al músculo, por lo que en las vesículas sinápticas hay muchas mitocondrias. Cuando un impulso nervioso llega a la unión neuromuscular, se liberan aproximadamente 125 vesículas de acetilcolina hacia el espacio sináptico, esto es posible ya que, al llegar el impulso, se abren los canales de calcio dependientes por voltaje, lo que hace que el calcio que está en la hendidura entre al terminal sináptico y que la acetilcolina salga del terminal a la hendidura mediante exocitosis.
Cuando es liberada, dos moléculas acetilcolina se unen a cada canal iónico dependiente de ligando ubicados en la fibra muscular, lo que provoca su apertura y la consiguiente entrada de sodio a la fibra muscular debido a la diferencia de gradientes de concentración. El diámetro del canal es de 0,65nm, tan grande como para que entren moléculas de calcio, sodio y potasio, cada canal puede transmitir de 15 a 30 mil iones sodio en 1ms, el tiempo que el canal permanece abierto es suficiente para excitar la fibra muscular, lo que eleva el potencial eléctrico de la misma hasta entre los 50 y los 75mV y a su vez generando lo que se denomina como potencial de la placa terminal
La acetilcolina es destruida por medio de dos medios, la mayor parte es destruida por la enzima acetilcolinesterasa, que se encuentra en la capa esponjosa de tejido conjuntivo del espacio sináptico, está la responsable de convertir la acetilcolina en iones acetato y colina, esta última puede ser reutilizada, lo que resta de la acetilcolina se difunde hacia el exterior y queda inutilizable para la membrana de la fibra muscular, la rápida eliminación de la acetilcolina impide la reexcitación muscular después de un potencial de acción. Una unión neuromuscular tiene un elevado factor de seguridad, que es aproximadamente tres veces mayor que el voltaje necesario para producir una contracción muscular, pero cuando una unión se estimula más de 100 veces por segundo por varios minutos, la cantidad de vesículas de acetilcolina que pueden liberarse disminuye, por lo que se dice que la unión llega a un estado de fatiga, efecto que también se aplica en la sinapsis en el sistema nervioso central.
Biología molecular de la formación y liberación de acetilcolina
Se forman aproximadamente 300,000 vesículas en el aparato de Golgi del cuerpo de la motoneurona de la médula espinal y se transporta a lo largo de todo el axoplasma hasta la unión neuromuscular, cada vesícula tiene un tamaño aproximado de 40nm.
La acetilcolina es sintetizada en el citosol de la terminación nerviosa y atraviesa inmediatamente la membrana de las vesículas, cada vesícula contendrá aproximadamente 10,000 moléculas de acetilcolina.
Cuando el potencial llega a la terminación nerviosa, se abren los canales de calcio, esta apertura aumenta la velocidad de fusión de las vesículas que transportan la acetilcolina con la membrana de la terminación, liberando la acetilcolina en la hendidura sináptica. Posteriormente se elimina la acetilcolina mediante los medios mencionados con anterioridad. El tiempo de esta secuencia de acontecimientos es de 5 a 10ms.
Las vesículas disponibles alcanzan para transmitir algunos miles de impulsos desde la fibra nerviosa hacia la muscular. Unos segundos después aparecen hendiduras revestidas en la membrana de la terminación nerviosa producidas especialmente por clatrina que, posteriormente, se rompen por la contracción de las mismas proteínas para formar nuevas vesículas. Unos segundos más tarde, la acetilcolina es transportada alas vesículas para una nueva liberación de acetilcolina.
Existen fármacos actúan de manera similar a la acetilcolina, tales como la metacolina, el carbacol y la nicotina. La diferencia entre estos y la acetilcolina es que no son destruidos por la colinesteraza o las destruyen tan lento que el efecto se queda de varios minutos a horas. Los tres fármacos crean zonas de despolarización en la membrana de la fibra muscular, lo que genera espasmos musculares debidos a la fuga de iones que inician un nuevo impulso nervioso. Otros fármacos, como la neostigmina y la fisostigmina, inactivan el proceso de la acetilcolinesterasa combinándose con ella, lo que hace que la acetilcolina se acumule con cada potencial de acción y producen espasmos musculares. En caso de que el espasmo sea laríngeo, se puede producir la muerte por asfixia. Después de varias horas los fármacos se separan de la acetilcolinesterasa y esta se activa de nuevo. Así como hay fármacos que potencian la transmisión por unión neuromotora, hay otros que la bloquean, tales como la d-tubocurarina, que bloquean la acción de la acetilcolina sobre sus receptores.
Miastenia grave
causa de debilidad muscular esta aparece en 1 de cada 20,000 personas y hace que las uniones neuromusculares no puedan transferir suficientes señales, por lo que la fibra muscular no se despolariza, lo que provoca debilidad muscular. La enfermedad se detecta la observación de anticuerpos frente a los receptores de acetilcolina, por lo que se infiere que la enfermedad crea dichos anticuerpos los cuales destruyen los receptores de acetilcolina en las membranas postsinápticas. Cuando la enfermedad es muy fuerte, el paciente puede morir por debilidad en los músculos respiratorios. Uno de los tratamientos contra la Miastenia grave es la administración de neostigmina.
Potencial de acción muscular El inicio y la conducción de los potenciales de acción muscular son similares a los del sistema nervioso en algunas cosas:
Su potencial de membrana en reposo es el mismo.
La duración del potencial de acción en las fibras del músculo esquelético es 5 veces mayor que en los nervios mielinizados grandes.
La velocidad de conducción es 1/13 de la velocidad de las fibras nerviosas mielinizadas grandes que excitan el músculo esquelético.
Los potenciales que se propagan en fibras musculares lo hacen generalmente por su superficie, en caso de necesitar una contracción muscular máxima, los impulsos se deben generar en lo más profundo del músculo penetrando en las miofibrillas, esto se logra gracias a los túbulos T que se originan en la membrana y atraviesan la fibra muscular perpendicularmente produciendo la liberación de calcio en su luz que a su vez producen la contracción. Esto se denomina excitacióncontracción.
Junto a los túbulos T se encuentran estructuras llamadas retículos sarcoplásmicos, por cavidades llamadas cisternas terminales y túbulos longitudinales que envuelven las miofibrillas. En el interior de estos últimos hay muchos iones calcio que son liberados debido a un potencial de acción en un túbulo T cercano.
El mecanismo funciona así: el túbulo T recibe el potencial, el voltaje es captado por receptores de dihidropiridina ligados a canales de liberación de calcio de las cisternas reticulares, esto abre los canales liberadores de calcio sobre el sarcoplasma y provoca la contracción del músculo. Después de iniciada la contracción, una bomba de calcio que está en el retículo sarcoplásmico bombea estos iones desde las miofibrillas a los túbulos sarcoplásmicos. Esta bomba puede concentrar 10,000 veces los iones calcio en los túbulos.
Capítulo 8: Excitación y contracción del músculo liso
El músculo liso, que está formado por fibras mucho menores, habitualmente de 1 a 5 μm de diámetro y de solo 20 a 500 μm de longitud. Las fibras musculares esqueléticas tienen un diámetro hasta 30 veces mayor y una longitud cientos de veces mayor, hay que tomar en cuenta que las mismas fuerzas de atracción entre los filamentos de miosina y actina producen la contracción en el músculo liso y en el músculo esquelético, pero la disposición física interna de las fibras musculares lisas es diferente. El músculo liso de los distintos órganos es distinto del de la mayor parte de los demás en varios sentidos: dimensiones físicas, organización en fascículos o láminas, respuesta a diferentes tipos de estímulos, características de la inervación, y función, pero se puede dividir en multiunitario y unitario.
El multiunitario está formado por fibras musculares lisas separadas y discretas. Cada una de las fibras actúa independientemente de las demás y con frecuencia está inervada por una única terminación nerviosa, como ocurre en las fibras musculares esqueléticas. Además, la superficie externa de estas fibras, como en el caso de las fibras musculares esqueléticas, está cubierta por una capa delgada de sustancia similar a una membrana basal, una mezcla de colágeno fino y glucoproteínas que aísla las fibras separadas entre sí.
El unitario se denomina músculo liso sincitial o músculo liso visceral. El término “unitario” es confuso porque no se refiere a fibras musculares únicas. Por el contrario, se refiere a una masa de cientos a miles de fibras musculares lisas que se contraen juntas como una única unidad.
El músculo liso contiene filamentos tanto de actina como de miosina, que tienen características químicas similares a los filamentos de actina y miosina del músculo esquelético. No contiene el complejo de troponina necesario para el control de la contracción del músculo esquelético, de modo que el mecanismo de control de la contracción es diferente. Los filamentos de actina y miosina del músculo liso interactúan entre sí de manera muy similar a como lo hacen en el músculo esquelético. Además, el proceso contráctil es activado por los iones calcio, y el trifosfato de adenosina (ATP) se degrada a difosfato de adenosina (ADP) para proporcionar la energía para la contracción. La Base física de la contracción del músculo liso muestra grandes números de filamentos de actina unidos a los cuerpos densos. Algunos de estos cuerpos están unidos a la membrana celular; otros están dispersos en el interior de la célula. Algunos de los cuerpos densos de la membrana de células adyacentes están unidos entre sí por puentes proteicos intercelulares.
La fuerza de contracción se transmite de unas células a otras principalmente a través de estos enlaces. La mayoría de los músculos esqueléticos se contraen y relajan rápidamente, muchas de las contracciones del músculo liso son contracciones tónicas prolongadas, que a veces duran horas o incluso días. Por tanto, cabe esperar que las características físicas y químicas de la contracción del músculo liso sean diferentes de las del músculo esquelético. La rapidez del ciclado de los puentes transversos de miosina en el músculo liso es mucho más lenta que en el músculo esquelético. A pesar de todo, se piensa que la fracción de tiempo que los puentes cruzados permanecen unidos a los filamentos de actina, que es un factor importante que determina la fuerza de la contracción, está muy aumentada en el músculo liso. Para mantener la misma tensión de contracción en el músculo liso que en el músculo esquelético solo es necesario de 1/10 a 1/300 de energía. También se piensa que esto se debe al lento ciclado de unión y separación de los puentes cruzados y a que solo es necesaria una molécula de ATP para cada ciclo, independientemente de su duración. Un tejido muscular liso típico comienza a contraerse de 50 a 100 ms después de ser excitado, alcanza la contracción completa aproximadamente 0,5 s después, y después la fuerza contráctil disminuye en 1 a 2 s más, dando un tiempo total de contracción de 1 a 3 s. Este tiempo es aproximadamente 30 veces más prolongado que una contracción única de una fibra muscular esquelética media. El inicio lento de la contracción del músculo liso, así como su contracción prolongada, está producido por la lentitud de la unión y la separación de los puentes cruzados a los filamentos de actina. La fuerza máxima de contracción del músculo liso es con frecuencia mayor que la del músculo esquelético, hasta 4 a 6 kg/cm2 de área transversal para el músculo liso, en comparación con 3 a 4 kg para el músculo esquelético.
La magnitud de la excitación continuada habitualmente se puede reducir a mucho menos del nivel inicial, a pesar de lo cual el músculo mantiene su fuerza de contracción completa. Además, la energía que se consume para mantener la contracción con frecuencia es minúscula, a veces tan solo 1/300 de la energía necesaria para una contracción sostenida y comparable del músculo esquelético. Este mecanismo se denomina efecto de cerrojo. Una vez que el músculo liso ha generado la contracción máxima, la magnitud de la excitación continuada habitualmente se puede reducir a mucho menos del nivel inicial, a pesar de lo cual el músculo mantiene su fuerza de contracción completa.
El mecanismo de cerrojo es que permite mantener una contracción tónica prolongada en el músculo liso durante horas con un bajo consumo de energía. Otra característica importante del músculo liso es su capacidad de recuperar casi su fuerza de contracción original segundos a minutos después de que haya sido alargado o acortado.
Regulación de la contracción por los iones calcio
El estímulo que inicia la mayoría de las contracciones del músculo liso es un aumento de los iones calcio en el medio intracelular. Este aumento puede estar producido en diferentes tipos de músculo liso por la estimulación nerviosa de las fibras de músculo liso, por estimulación hormonal, por distensión de la fibra o incluso por cambios del ambiente químico de la fibra. En lugar de la troponina, las células musculares lisas contienen una gran cantidad de otra proteína reguladora denominada calmodulina.
La calmodulina inicia la contracción al activar los puentes cruzados de miosina. Esta activación y la posterior contracción se producen según la siguiente secuencia:
La concentración de calcio en el líquido citosólico del músculo liso se incrementa como consecuencia de la entrada de calcio desde el líquido extracelular a través de los canales de calcio y/o la liberación de calcio desde el retículo sarcoplásmico.
Los iones calcio se unen a la calmodulina de forma reversible. El complejo calmodulina-calcio se une después a la miosina cinasa de cadena ligera, que es una enzima fosforiladora, y la activa.
Una de las cadenas ligeras de cada una de las cabezas de miosina, denominada cabeza reguladora, se fosforila en respuesta a esta miosina cinasa.
Cuando esta cadena no está fosforilada no se produce el ciclo de unión-separación de la cabeza de miosina con el filamento de actina. Sin embargo, cuando la cadena reguladora está fosforilada la cabeza tiene la capacidad de unirse repetitivamente al filamento de actina y de avanzar a través de todo el proceso de ciclado de «tirones» intermitentes, al igual que ocurre en el músculo esquelético, produciendo de esta manera la contracción muscular. El retículo sarcoplásmico, que proporciona prácticamente todos los iones calcio para la contracción musculoesquelética, está desarrollado solo ligeramente en la mayor parte del músculo liso. En su lugar, la mayoría de los iones calcio que provocan la contracción entran en la célula muscular desde el líquido extracelular en el momento del potencial de acción u otro estímulo. Es decir, la concentración de iones calcio en el líquido extracelular es superior a 10−3 molar, en comparación con un valor inferior a 10−7 molar en el interior de la célula muscular lisa; esta situación origina una rápida difusión de los iones calcio en la célula desde el líquido extracelular cuando se abren los canales de calcio. Los retículos sarcoplásmicos muy poco desarrollados que se sitúan cerca de las membranas celulares en algunas células del músculo liso más grandes. Pequeñas invaginaciones de la membrana celular, denominadas cavéolas, terminan en las superficies de estos túbulos. Las cavéolas sugieren una rudimentaria analogía del sistema de túbulos transversos del músculo esquelético.
Cuando se transmite un potencial de acción a las cavéolas, según se cree estimula la liberación de iones calcio desde los túbulos sarcoplásmicos contiguos de la misma forma que los potenciales de acción en los túbulos transversos del músculo esquelético provocan la liberación de iones calcio desde los túbulos sarcoplásmicos transversos. . Cuando la concentración extracelular de iones calcio disminuye a aproximadamente entre 1/3 y 1/10 de la normal, la contracción del músculo liso suele cesar. Por tanto, la fuerza de la contracción del músculo liso suele ser muy dependiente de la concentración de los iones calcio en el líquido extracelular. Para provocar la relajación del músculo liso después de que se haya contraído es preciso extraer los iones calcio de los líquidos intracelulares. Esta extracción se consigue mediante una bomba de calcio que bombea los iones calcio fuera de la fibra de músculo liso de nuevo al líquido extracelular, o al retículo sarcoplásmico, si estuviera presente. La relajación del músculo liso tiene lugar cuando los canales de calcio se cierran y la bomba de calcio transporta iones calcio fuera del líquido citosólico de la célula. Cuando la concentración de iones calcio disminuye por debajo de un nivel crítico, los procesos que se acaban de señalar se invierten automáticamente, excepto la fosforilación de la cabeza de miosina.
La inversión de esta reacción precisa otra enzima, la miosina fosfatasa, que está localizada en el citosol de la célula muscular lisa y que escinde el fosfato de la cadena ligera reguladora. Debido a la importancia del fenómeno de cerrojo en el músculo liso, y como este fenómeno permite el mantenimiento a largo plazo del tono en muchos órganos que tienen músculo liso sin un gran gasto de energía, se han hecho muchos intentos de explicarlo. Entre los muchos mecanismos que se han propuesto, uno de los más sencillos es el siguiente. Cuando las enzimas miosina cinasa y miosina fosfatasa están intensamente activadas, la frecuencia de ciclado de las cabezas de miosina y la velocidad de contracción son elevadas. Después, cuando disminuye la activación de las enzimas, lo hace también la frecuencia de ciclado, pero al mismo tiempo la desactivación de estas enzimas permite que las cabezas de miosina permanezcan unidas al filamento de actina durante una proporción cada vez mayor del período de ciclado. Por tanto, el número de cabezas unidas al filamento de actina en cualquier momento dado sigue siendo grande. Como el número de cabezas unidas a la actina determina la fuerza estática de la contracción, se mantiene, o «cierra», la tensión; sin embargo, el músculo utiliza poca energía porque el ATP no se degrada a ADP excepto en las pocas ocasiones en las que una cabeza se separa.
El cuerpo humano está formado con el 40% de musculo esquelético y el 10% de musculo liso y cardíaco. Los músculos esqueléticos tienen como principal función dar movimiento al esqueleto. Los músculos esqueléticos están formados por numerosas fibras que se extienden a los largo de la longitud de los músculos y están inervadas por una sola terminación nerviosa cerca del punto medio de la fibra.
Las fibras del musculo esquelético se encuentran envueltas por el sarcolema que está formado por una membrana plasmática y una cubierta externa de polisacáridos que contiene fibrillas de colágeno. La capa superficial del sarcolema se fusiona con fibras tendinosas que se agrupan y forman tendones los cuales se encargan de insertar los músculos a los huesos. Cada fibra muscular está compuesta por miofibrillas, y cada miofibrilla está compuesta por proteínas de actina y miosina que son las encargadas de la contracción muscular. Estos filamentos de actina y miosina se encuentran unidos por la proteína titina que se caracteriza por ser elástica, actúan como armazón de estas dos proteínas y sirve de molde de formación inicial de porciones de los filamentos contráctiles del sarcomero.
La actina forma bandas claras (bandas I) son isotropas y la miosina forma bandas oscuras (bandas A) son anisótropas. Estas bandas claras y oscuras dan al musculo esquelético el aspecto estriado. Los puentes cruzados son los extremos de los filamentos de miosina y a los extremos de los filamentos de actina se les llama disco Z, y por último, la porción que se encuentra entre los discos Z se llama sarcomero. Las miofibrillas contienen un líquido llamado sarcoplasma entre sus espacios, este líquido está formado por potasio, magnesio, fosfato, enzimas proteicas y mitocondrias que tienen una función muy importante ya que le brindan energía a las miofibrillas en contracción en forma de ATP.
Otro elemento importante de las miofibrillas es el retículo sarcoplasmico, este se encarga de regular el almacenamiento, la liberación y recaptación del calcio para controlar la contracción muscular.
MECANISMO GENERAL DE LA CONTRACCIÓN MUSCULAR
•Un potencial de acción viaja a lo largo de una fibra motora hasta sus terminales sobre las fibras musculares
•En cada terminal, el nervio secreta una pequeña cantidad de la sustancia neurotransmisora acetilcolina.
•La acetilcolina actúa en una zona local de la membrana de la fibra muscular para abrir múltiples canales de cationes «activados por acetilcolina» a través de moléculas proteicas que flotan en la membra
•. La apertura de los canales activados por acetilcolina permite que grandes cantidades de iones sodio difundan hacia el interior de la membrana de la fibra muscular. Esta acción provoca una despolarización local que, a su vez, conduce a la apertura de los canales de sodio activados por el voltaje, que inicia un potencial de acción en la membrana.
•El potencial de acción viaja a lo largo de la membrana de la fibra muscular de la misma manera que los potenciales de acción viajan a lo largo de las membranas de las fibras nerviosas.
•El potencial de acción despolariza la membrana muscular, y buena parte de la electricidad del potencial de acción fluye a través del centro de la fibra muscular, donde hace que el retículo sarcoplásmico libere grandes cantidades de iones calcio que se han almacenado en el interior de este retículo.
•Los iones calcio inician fuerzas de atracción entre los filamentos de actina y miosina, haciendo que se deslicen unos sobre otros en sentido longitudinal, lo que constituye el proceso contráctil.
•Después de una fracción de segundo los iones calcio son bombeados de nuevo hacia el retículo sarcoplásmico por una bomba de Ca++ de la membrana y permanecen almacenados en el retículo hasta que llega un nuevo potencial de acción muscular; esta retirada de los iones calcio desde las miofibrillas hace que cese la contracción muscular.
MECANISMO MOLECULAR DE LA CONTRACCIÓN MUSCULAR
El deslizamiento de los filamentos produce la contracción muscular, este mecanismo se lleva a cabo de la siguiente manera: en el estado contraído los filamentos de actina se deslizan entre los filamentos de miosina esta acción se lleva a cabo por las fuerzas generadas entre los puentes cruzados que se encuentran en ambos filamentos. Cuando un potencial de acción viaja a lo largo de la fibra muscular provoca la liberación de iones de calcio del retículo sarcoplasmico que rodean a las miofibrillas y al mismo tiempo estas fuerzas de atracción entre los filamentos y comienza la contracción.
CARACTERÍSTICAS MOLECULARES DE LOS FILAMENTOS CONTRÁCTILES
Los filamentos de miosina están formados por 200 o más moléculas de miosina. Las moléculas de miosina están formadas por seis cadenas polipeptidicas, dos cadenas pesadas y cuatro cadenas ligeras.
Los filamentos de actina están formados por la molécula de la proteína F-actina bicantenaria, tropiomiosina y troponina. La F-actina esta presentada por dos hebras de color claro, latropiomiosina se encuentra enrollada en espiralalrededor de los lados de la hélice de F-actina y por último, la troponina se ubica a los lados de las moléculas tropiomiosina, la troponina se divide en tres subunidades: troponina T, troponina C y troponina I. Cuando ocurre la inhibición del filamento de actina puro sin la presencia del complejo troponina-tropomiosina los puntos activos del filamento de actina normal del musculo relajado son inhibidos físicamente por el complejo troponinatropoiosina.
En presencia de iones de calcio, se inhibe el propio efecto inhibidor del complejo troponinatropiomiosina sobre los filamentos de actina. Una vez activado el filamento de actina por los iones de calcio la cabeza de los puentes cruzados de los filamentos de miosina son atraídos hacia los puntos activos del filamento de actina y produce la contracción. En la contracción se utilizan grandes cantidades de energía de ATP para formar ADP. La longitud del sarcomero varía dependiendo de la cantidad de superposición entre los filamentos de miosina y actina durante la tensión activa. -la tensión activa (aumento de la tensión que se produce durante la contracción) se reduce a medida que el musculo es distendido más allá de su longitud normal.
ATP como fuente de energía para la contracción: fenómenos químicos en el movimiento de las cabezas de miosina.
Antes de que comience la contracción, las cabezas de los puentes cruzados se unen al ATP. La actividad ATPasa de la cabeza de miosina escinde inmediatamente el ATP, aunque deja los productos de la escisión, el ADP y el ion fosfato, unidos a la cabeza. En este estado la conformación de la cabeza es tal que se extiende perpendicularmente hacia el filamento de actina, pero todavía no está unida a ella.
Cuando el complejo troponina-tropomiosina se une a los iones calcio quedan al descubierto los puntos activos del filamento de actina, y entonces las cabezas de miosina se unen a ello
El enlace entre la cabeza del puente cruzado y el punto activo del filamento de actina produce un cambio conformacional de la cabeza, lo que hace que la cabeza se desplace hacia el brazo del puente cruzado. Esto proporciona el golpe activo para tirar del filamento de actina. La energía que activa el golpe activo es la energía que ya se ha almacenado, como un muelle «comprimido» por el cambio conformacional que se había producido previamente en la cabeza cuando se escindió la molécula de ATP.
Una vez que se desplaza la cabeza del puente cruzado, esto permite la liberación del ADP y el ion fosfato que previamente estaban unidos a la cabeza. En el punto de liberación del ADP se une una nueva molécula de ATP. Esta unión de una nueva molécula de ATP hace que la cabeza se separe de la actina.
Después de que la cabeza se haya separado de la actina, se escinde la nueva molécula de ATP para comenzar el ciclo siguiente, dando lugar a un nuevo golpe activo. Es decir, la energía una vez más «comprime» la cabeza de nuevo a su situación perpendicular, dispuesta para comenzar el nuevo ciclo de golpe activo.
Cuando la cabeza comprimida (con su energía almacenada procedente del ATP escindido) se une a un nuevo punto activo del filamento de actina, se estira y una vez más proporciona un nuevo golpe activo. Figura
ENERGÉTICA DE LA CONTRACCIÓN MUSCULAR La energía que se necesita para realizar un trabajo proviene de las reacciones químicas de las células musculares durante la contracción. Existen tres fuentes de energía importantes utilizadas para reconstruir el ATP: fosfocreatina, glucolisis y metabolismo oxidativo.
CARACTERÍSTICAS DE LA CONTRACCIÓN DE TODO EL MUSCULO
Las contracciones musculares se caracterizan por ser isométricas e isotónicas. Las contracciones isométricas son todas aquellas en las cuales al ejercer una fuerza el musculo se contrae y se relaja sin afectar su longitud, y en las contracciones isotónicas los músculos se acortan contra una carga fija.
Los músculos existen en diferentes tamaños, desde el más pequeños hasta el más grande, con fibras cortas y largas, es por eso que la contracción de cada uno de ellos es variable. Las fibras que inervan los musculos se clasifican en rapidas y lentas: 1) Fibras lentas (tipo I, musculo rojo) • Pequeñas • Inervadas por fibras nerviosas pequeñas • Sistema de vascularización extenso • Número elevado de mitocondrias • Grandes cantidades de mioglobina 2) Fibras rápidas (tipo II, musculo blanco) • Grandes • Retículo sarcoplasmico extenso • Enzimas glucoliticas • Vascularización menos extensa • Menos mitocondrias • Déficit de mioglobina
MECÁNICA DE LA CONTRACCIÓN DEL MUSCULO ESQUELÉTICO
Unidad motora: Son todas las fibras musculares inervadas por una única fibra nerviosa. Los músculos pequeños tienen un movimiento rápido y específico por lo que están inervadas por más fibras nerviosas y menos fibras musculares, de lo contrario los músculos grandes que no tienen un control fino tienen más fibras musculares en una unidad motora. La sumación de las fuerzas musculares se distingue en dos tipos:
1) Sumacion de fibras múltiples: aumenta el número de unidades motoras que se contraen de manera simultánea. 2) Sumacion de frecuencia y tetanizacion: aumenta la frecuencia de la contracción sin permitir una relajación entre los potenciales de acción. La máxima fuerza de contracción en un musculo en función normal es de 3 y 4 kg por centímetro cuadrado de musculo. Efecto escalera o treppe se realiza cuando un musculo empieza a contraerse después de un periodo de reposo prolongado, su fuerza inicial de contracción puede ser muy pequeña y va aumentando hasta una meseta. Esto es posible gracias al aumento de iones calcio en el citosol debido a la liberación de más iones en el retículo sarcoplasmico. La fatiga muscular es la contracción larga e intensa de un musculo, esta aumenta de acuerdo a la velocidad de depleción de glucógeno en el musculo. La cinesiología se encarga del estudio de los diferentes tipos de músculos y sistemas de palanca. Los sistemas de palanca están formados por huesos, los cuales tienen una tensión aplicada en sus puntos de inserción. Estos se encargan del movimiento y la intensidad en la que se realiza. La coactivación es un proceso de contracción simultanea de músculos agonistas y antagonistas de lados opuestos de las articulaciones controlados por los centros de control del encéfalo y de la medula espinal.
REMODELACIÓN DEL MUSCULO PARA ADAPTARSE A LA FUNCIÓN
Los músculos se modelan frecuentemente para adaptarse a sus funciones, alteran su longitud, diámetro, fuerza, vascularización y muy levemente se modelan sus tipos de fibras musculares. Los músculos tienden a obtener mayor volumen o bien la disminución de este, son denominados hipertrofia muscular y atrofia muscular respectivamente. La hipertrofia muscular se debe al aumento de filamentos de actina y miosina en cada fibra muscular, también existe un aumento en los sistemas enzimáticos que proporcionan energía. La atrofia muscular es la degradación proteica del musculo importada por la ruta de ubicuitina-proteasoma dependiente del ATP. Cuando un musculo pierde su inervación ya no recibe las señales contráctiles que son necesarias para mantener el tamaño muscular normal, por lo que comienzan a haber cambio degenerativo en las fibras musculares, esta es destruido y sustituido por tejido fibroso y adiposo. Rigidez cadavérica: como su nombre lo dice, algunas horas después de la muerte los músculos del cuerpo entran en un estado de resistencia, se contraen, esto se debe a la perdida de ATP que es el encargado de la separación de puentes cruzados de los filamentos de actina. La distrofia: muscular es la debilidad y degeneración progresiva de las fibras musculares.
Hay potenciales eléctricos a través de las membranas de prácticamente
todas las células del cuerpo. Además, algunas células, como las células
nerviosas y musculares, son capaces de generar impulsos electroquímicos
rápidamente cambiantes en sus
membranas, y estos impulsos se utilizan para transmitir señales a través de las
membranas de los nervios y de los músculos.
En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células. Este análisis se refiere a los potenciales de membrana que se generan tanto en reposo como durante la acción en las células nerviosas y musculares.
Física básica de los potenciales de membrana
Potenciales de membrana provocados por difusión
«Potencial de difusión»
producido por una diferencia de concentración iónica a los dos lados de
la membrana. Debido al gran gradiente de concentración de
potasio desde el interior hacia el exterior hay una intensa tendencia a que
cantidades adicionales de iones potasio difundan hacia fuera a través de la
membrana.
En un plazo de aproximadamente 1 ms la diferencia de potencial entre el
interior y el exterior, denominada potencial de difusión, se hace lo
suficientemente grande como para bloquear la difusión adicional neta de potasio
hacia el exterior, a pesar del elevado gradiente de concentración iónica de
potasio. En la fibra nerviosa normal del mamífero la diferencia de potencial
necesaria es de aproximadamente 94 mV, con negatividad en el interior de la
membrana de la fibra.
La difusión de los iones sodio de carga positiva hacia el interior crea
un potencial de membrana de polaridad opuesta, con negatividad en el
exterior y positividad en el interior. Una vez más el potencial de membrana se
hace lo suficientemente elevado en un plazo de milisegundos como para bloquear
la ulterior difusión neta de iones sodio hacia el interior; sin embargo, esta
vez, en la fibra nerviosa del mamífero, el potencial es de aproximadamente
61 mV positivos en el interior de la fibra.
Relación del potencial de difusión con la diferencia de concentración:
potencial de Nernst.
El nivel del potencial de difusión a través de una membrana que se opone
exactamente a la difusión neta de un ion particular a través de la membrana se
denomina potencial de Nernst.
La magnitud de este potencial de Nernst viene determinada por el cociente
de las concentraciones de ese ion específico en los dos lados de la
membrana. Cuanto mayor es este cociente, mayor es la tendencia del ion a
difundir en una dirección y, por tanto, mayor será el potencial de Nernst
necesario para impedir la difusión neta adicional.
Se calcula que el potencial de Nernst es de –61 mV en el interior de la membrana.
Medición del potencial de membrana
El método para medir el potencial de membrana es simple en teoría,
aunque con frecuencia es difícil en la práctica debido al pequeño tamaño de la
mayor parte de las fibras. La pipeta se inserta en la membrana celular hasta el
interior de la fibra. Después se coloca otro electrodo, denominado «electrodo
indiferente», en el líquido extracelular, y se mide la diferencia de potencial
entre el interior y exterior de la fibra utilizando un voltímetro adecuado.
Se debe transferir un número increíblemente pequeño de iones a través de
la membrana para establecer el «potencial en reposo» normal de –90mV en el
interior de la fibra nerviosa; esto significa que sólo se debe transferir entre
1/3.000.000 a 1/100.000.000 del número total de cargas positivas del interior
de la fibra. Además, un número igual de pequeño de iones positivos que se
mueven desde el exterior hacia el interior de la fibra puede invertir el
potencial desde –90mV hasta tanto
como +35mV en tan sólo 1/10.000 de
segundo.
Potencial de membrana en reposo de los nervios
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90mV. Es decir, el potencial en el interior de la fibra es 90mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.
Transporte activo de los iones sodio y potasio a través de la membrana:
la bomba sodio-potasio (Na+-K+).
Se trata de una bomba electrógena porque se bombean más cargas
positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por
cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto
genera un potencial negativo en el interior de la membrana celular.
La bomba Na+-K+ también genera grandes gradientes de concentración para el sodio y el
potasio a través de la membrana nerviosa en reposo. Estos gradientes son los
siguientes:
Na+ (exterior):
142 mEq/lNa+ (interior):
14 mEq/l
K+ (exterior):
4 mEq/lK+ (interior):
140 mEq/l
Fuga de potasio y de sodio a través de la membrana nerviosa.
«dominio de poros en tándem», canal de potasio o canal de «fuga» de potasio (K+), en la membrana nerviosa a través de la que pueden escapar iones potasio incluso en una célula en reposo.
Origen del potencial de membrana en reposo normal
Contribución de la difusión de sodio a través de la membrana nerviosa.
Esto se puede responder utilizando la ecuación de Goldman que se ha
descrito previamente. Intuitivamente se puede ver que, si la membrana es muy
permeable al potasio pero sólo ligeramente permeable al sodio, es lógico que la
difusión del potasio contribuya mucho más al potencial de membrana que la
difusión del sodio. En la fibra nerviosa normal la permeabilidad de la membrana
al potasio es aproximadamente 100 veces mayor que la permeabilidad al sodio.
Utilizando este valor en la ecuación de Goldman se obtiene un potencial en el
interior de la membrana de –86mV,
Contribución de la bomba Na+-K+.
El hecho de que se bombeen más iones sodio hacia el exterior que iones
potasio hacia el interior da lugar a una pérdida continua de cargas positivas
desde el interior de la membrana; esto genera un grado adicional de negatividad
(aproximadamente –4mV más) en el interior además del que se puede explicar por
la difusión de manera aislada. Por tanto, el potencial de membrana neto
cuando actúan todos estos mecanismos a la vez es de aproximadamente –90 mV.
En resumen, los potenciales de difusión aislados que pro- duce la difusión del sodio y del potasio darían un potencial de membrana de aproximadamente –86 mV, casi todo determinado por la difusión de potasio. Además, se generan –4mV adicionales al potencial de membrana por la acción continua de la bomba de Na+-K+ electrógena, generándose un poten- cial neto de membrana de –90 mV.
Potencial de acción
nervioso
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega al extremo de la misma.
Las sucesivas fases del potencial de
acción son las siguientes.
Fase de reposo. Este es
el potencial de membrana en reposo antes del comienzo del potencial de acción.
Se dice que la membrana está «polarizada» durante esta fase debido al
potencial de membrana negativo de –90mV que está presente.
Fase de despolarización. En este momento
la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite
que un gran número de iones sodio con carga positiva difunda hacia el interior
del axón. El estado «polarizado» normal de –90mV se neutraliza inmediatamente
por la entrada de iones sodio cargados positivamente, y el potencial aumenta
rápidamente en dirección positiva. Esto se denomina despolarización.
Fase de repolarización. En un plazo de
algunas diezmilésimas de segundo después de que la membrana se haya hecho muy
permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los
canales de potasio se abren más de lo normal. De esta manera, la rápida
difusión de los iones potasio hacia el exterior restablece el potencial de membrana
en reposo negativo normal. Esto se denomina repolarización de la
membrana.
Canales de sodio y potasio activados por el voltaje
El actor necesario en la producción tanto de la despolarización como de
la repolarización de la membrana nerviosa durante el potencial de acción es el canal
de sodio activado por el voltaje. Un canal de potasio activado por el
voltaje también tiene una función importante en el aumento de la rapidez de
la repolarización de la membrana. Estos dos cana- les activados por el voltaje
tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga K+.
Canal de sodio activado por el voltaje: activación e inactivación del
canal
Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. La parte superior izquierda de la figura representa el estado de estas dos compuertas en la membrana en reposo normal, cuando el potencial de membrana es de –90 mV. En este estado la compuerta de activación está cerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra a través de estos canales de sodio.
Activación del canal de sodio. Cuando
el potencial de membrana se hace menos negativo que durante el estado de
reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje
(habitualmente algún punto entre –70 y –50mV) que produce un cambio
conformacional súbito en la activación de la compuerta, que bascula totalmente
hasta la posición de abierta. Esto se denomina estado activado; durante
este estado los iones sodio pueden atravesar el canal, aumentando la
permeabilidad de la membrana al sodio hasta 500 a 5.000 veces.
Inactivación del canal de sodio.
El mismo aumento de voltaje que abre la compuerta de activación también cierra
la compuerta de inactivación. Sin embargo, la compuerta de inactivación se
cierra algunas diezmilésimas de segundo después de que se abra la compuerta de
activación. Es decir, el cambio conformacional que hace bascular la compuerta
de inactivación hacia el estado cerrado es un proceso algo más lento que el
cambio conformacional que abre la compuerta de activación.
Canal de potasio activado por el voltaje y su activación
Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90mV hacia cero, este voltaje produce una apertura conformacional de la compuerta y permite el aumento de la difusión de potasio hacia fuera a través del canal. Sin embargo, debido a la ligera demora de la apertura de los canales de potasio, en su mayor parte, se abren al mismo tiempo que están comenzando a cerrarse los canales de sodio debido a su inactivación. Por tanto, la disminución de la entrada de sodio hacia la célula y el aumento simultáneo de la salida de potasio desde la célula se combinan para acelerar el proceso de repolarización, lo que da lugar a la recuperación completa del potencial de membrana en reposo en otras pocas diezmilésimas de segundo.
Funciones de otros iones durante el potencial de acción
Hasta
ahora hemos considerado sólo la función de los iones sodio y potasio en la generación
del potencial de acción. Se deben considerar al menos otros dos tipos de iones:
los aniones negativos y los iones calcio.
Iones
con carga negativa (aniones) no difusibles en el interior del axón nervioso. En el
interior del axón hay muchos iones de carga negativa que no pueden atravesar
los canales de la membrana. Incluyen los aniones de las moléculas proteicas y
de muchos compuestos de fosfato orgánicos, compuestos de sulfato y similares.
Por tanto, estos iones negativos no difusibles son responsables de la carga
negativa en el interior de la fibra cuando hay un déficit neto de iones potasio
de carga positiva y de otros iones positivos.
Iones
calcio. Las membranas de casi todas las células del cuerpo
tienen una bomba de calcio similar a la bomba de sodio, y el calcio coopera con
el sodio (o actúa en su lugar) en algunas células para producir la mayor parte
del potencial de acción. Esto deja una concentración celular interna de iones
calcio de aproximadamente 10–7 molar, en comparación con una concentración
externa de aproximadamente 10–3 molar.
El
probable mecanismo mediante el cual los iones calcio afectan a los canales de
sodio es el siguiente: estos iones parecen unirse a la superficie externa de la
molécula de la proteína del canal de sodio. Las cargas positivas de estos iones
calcio, a su vez, alteran el estado eléctrico de la propia proteína del canal
de sodio, lo que modifica el nivel de voltaje necesario para abrir la compuerta
de sodio.
Inicio del potencial de acción
Un ciclo de retroalimentación positiva abre los canales de sodio.
Primero,
siempre que no haya alteraciones de la membrana de la fibra nerviosa, no se produce
ningún potencial de acción en el nervio normal. Sin embargo, si algún episodio
produce una elevación suficiente del potencial de membrana desde –90 mV hacia
el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos
canales de sodio activados por el voltaje.
Esto per- mite la entrada rápida de iones sodio, lo que produce una
elevación adicional del potencial de membrana y abre aún más canales de sodio
activados por el voltaje y permite que se produzca una mayor entrada de iones
sodio hacia el interior de la fibra.
Umbral para el inicio del potencial de acción.
No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva que se ha descrito en el párrafo anterior. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Por tanto, un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90mV hasta aproximadamente –65mV habitualmente da lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.
Propagación del potencial de acción
En los párrafos anteriores hemos analizado el potencial de acción que se
produce en un punto de la membrana. Sin embargo, un potencial de acción que se
desencadena en cualquier punto de una membrana excitable habitualmente excita
porciones adyacentes de la membrana, dando lugar a la propagación del potencial
de acción a lo largo de la membrana. De esta manera el proceso de
despolarización viaja a lo largo de toda la longitud de la fibra. Esta
transmisión del proceso de despolarización a lo largo de una fibra nerviosa
muscular se denomina impulso nervioso
o muscular.
Principio del todo o nada.
Una vez que se ha originado un potencial de acción en cualquier punto de
la membrana de una fibra normal, el proceso de despolarización viaja por toda
la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no
lo son. Esto se denomina principio del todo o nada y se aplica a todos
los tejidos excitables normales.
Restablecimiento de los gradientes iónicos de sodio y potasio tras
completarselos potenciales de acción: la importancia del metabolismo de la energía
La propagación de cada potencial de acción a lo largo de una fibra
nerviosa reduce ligeramente las diferencias de concentración de sodio y de
potasio en el interior y en el exterior de la membrana, porque los iones sodio
difunden hacia el interior durante la despolarización y los iones potasio
difunden hacia el exterior durante la repolarización. Para un único potencial
de acción este efecto es tan pequeño que no se puede medir. Aun así, con el
tiempo se hace necesario restablecer las diferencias de las concentraciones de
membrana de sodio y de potasio. Esto se consigue por la acción de la bomba Na+-K+
Una característica especial de la bomba Na+-K+-ATPasa es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular. De hecho, la actividad de bombeo aumenta aproximadamente en proporción a la tercera potencia de esta concentración intracelular de sodio.
Meseta en algunos potenciales de acción
En algunos casos la membrana excitada no se repolariza inmediatamente
después de la despolarización; por el contrario, el potencial permanece en una
meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y
sólo después comienza la repolarización.
La causa de la meseta es una combinación de varios factores. En primer
lugar, en el proceso de despolarización del músculo cardíaco participan dos
tipos de canales: 1) los canales de sodio habituales activados por el voltaje,
denominados canales rápidos, y 2) los canales de calcio-sodio activados
por el voltaje, que tienen una apertura lenta y que, por tanto, se denominan canales
lentos.
Ritmicidad de algunos tejidos excitables: descarga repetitiva
Las descargas repetitivas autoinducidas aparecen normalmente en el
corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso
central. Estas descargas rítmicas producen: 1) el latido rítmico del corazón;
2) el peristaltismo rítmico de los intestinos, y 3) fenómenos neuronales, como
el control rítmico de la respiración.
Proceso de reexcitación necesario para la ritmicidad espontánea.
Para que se produzca ritmicidad espontánea la membrana, incluso en su
estado natural, debe ser lo suficientemente permeable a los iones sodio (o a
los iones calcio y sodio a través de los canales lentos de calcio- sodio) como
para permitir la despolarización automática de la membrana. Después de otra
demora de milisegundos o segundos la excitabilidad espontánea produce una nueva
despolarización y se produce espontáneamente un nuevo potencial de acción. Este
ciclo continúa de manera indefinida y produce la excitación rítmica
autoinducida del tejido excitable.
El flujo aumentado de salida de iones potasio desplaza grandes
cantidades de cargas positivas hacia el exterior de la membrana, dejando en el
interior de la fibra una negatividad mucho mayor de lo que se produciría de
otra manera. Esto continúa durante aproximadamente un segundo después de que
haya finalizado el potencial de acción anterior, acercando de esta manera el
potencial de membrana al potencial de Nernst del potasio. Este es un estado
denominado hiperpolarización
Características especiales de la transmisión de señales en los troncos nerviosos
Fibras
nerviosas mielinizadas y no mielinizadas.
.
Las fibras grandes son mielinizadas y las pequeñas no mielinizadas.
Un tronco nervioso medio contiene aproximadamente el doble de fibras no
mielinizadas que mielinizadas.que con frecuencia es mucho más gruesa
que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la
vaina de mielina hay un nódulo de Ranvier.
Las
células de Schwann depositan la vaina de mielina alrededor del axón de la
siguiente manera: en primer lugar, la membrana de una célula de Schwann rodea
el axón. Después, la célula de Schwann rota muchas veces alrededor del axón,
depositando múltiples capas de membrana de la célula de Schwann que contiene la
sustancia lipídica esfingomielina. Esta sustancia es un excelente
aislante eléctrico que disminuye el flujo iónico a través de la membrana
aproximadamente 5.000 veces. En la unión entre dos células de Schwann sucesivas
a lo largo del axón permanece una pequeña zona no aislada de sólo 2 a 3m de longitud
en la que los iones pueden seguir fluyendo con facilidad a través de la
membrana del axón entre el líquido extracelular y el líquido intracelular del
interior del axón. Esta zona se denomina nódulo de Ranvier.
Conducción
«saltatoria» en las fibras mielinizadas de un nódulo a otro.
Aunque casi no pueden fluir iones a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos; esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro. Así, el impulso nervioso recorre a saltos la fibra, lo que es el origen del término «saltatoria».
Velocidad
de conducción en las fibras nerviosas.
La
velocidad de conducción del potencial de acción en las fibras nerviosas varía
desde tan sólo 0,25m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la
longitud de un campo de fútbol en un segundo) en las fibras mielinizadas
grandes.
Excitación: el proceso de generación del potencial de acción
Básicamente,
cualquier factor que haga que los iones sodio comiencen a difundir hacia el
interior a través de la membrana en un número suficiente puede desencadenar la
apertura regenerativa automática de los canales de sodio. Esto se puede deber a
un trastorno mecánico de la membrana, a los efectos químicos sobre
la membrana o al paso de electricidad a través de la membrana.
Excitación
de una fibra nerviosa por un electrodo metálico cargado negativamente.
El
método habitual para excitar un nervio o un músculo en el laboratorio experimental
es aplicar electricidad a la superficie del nervio del músculo mediante dos
electrodos pequeños, uno de los cuales tiene carga negativa y el otro positiva.
Cuando se hace esto la membrana excitable se estimula en el electrodo negativo.
Umbral
de excitación y «potenciales locales agudos».
Un estímulo eléctrico negativo débil puede no ser capaz de excitar una fibra. Sin embargo, cuando aumenta el voltaje del estímulo se llega a un punto en el que se produce la excitación. Estos cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generar un potencial de acción, se denominan potenciales subliminales agudos.
«Período
refractario» tras un potencial de acción, durante el cual no se puede generar
un nuevo estímulo
No se
puede producir un nuevo potencial de acción en una fibra excitable mientras la
membrana siga despolarizada por el potencial de acción precedente. El motivo de
esto es que poco después del inicio del potencial de acción se inactivan los
canales de sodio (o los canales de potasio, o ambos), y ninguna magnitud de la
señal excitadora que se aplique a estos canales en este momento abrirá las
compuertas de inactivación.
La
única situación que permitirá que se vuelvan a abrir es que el potencial de
membrana vuelva al nivel del potencial de membrana en reposo original o cerca
del mismo. Entonces, en otra pequeña fracción de segundo se abren las
compuertas de inactivación del canal y se puede iniciar un nuevo potencial de
acción.
Inhibición
de la excitabilidad: «estabilizadores»y
anestésicos locales
Al
contrario de los factores que aumentan la estabilidad nerviosa, otros factores,
denominados factores estabilizadores de la membrana, pueden reducir
la excitabilidad. Por ejemplo, una concentración elevada de calcio en el
líquido extracelular reduce la permeabilidad de la membrana a los iones
sodio y reduce simultáneamente la excitabilidad. Por tanto, se dice que el ion
calcio es un «estabilizador».






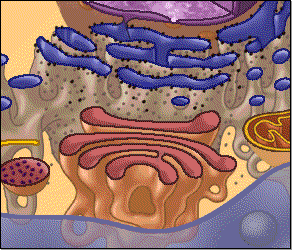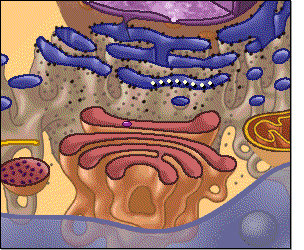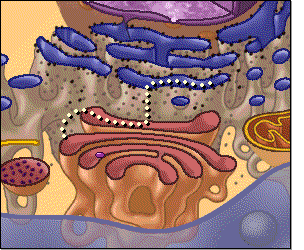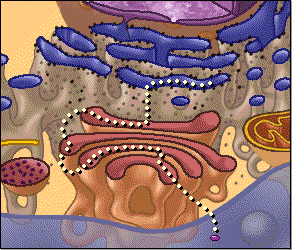




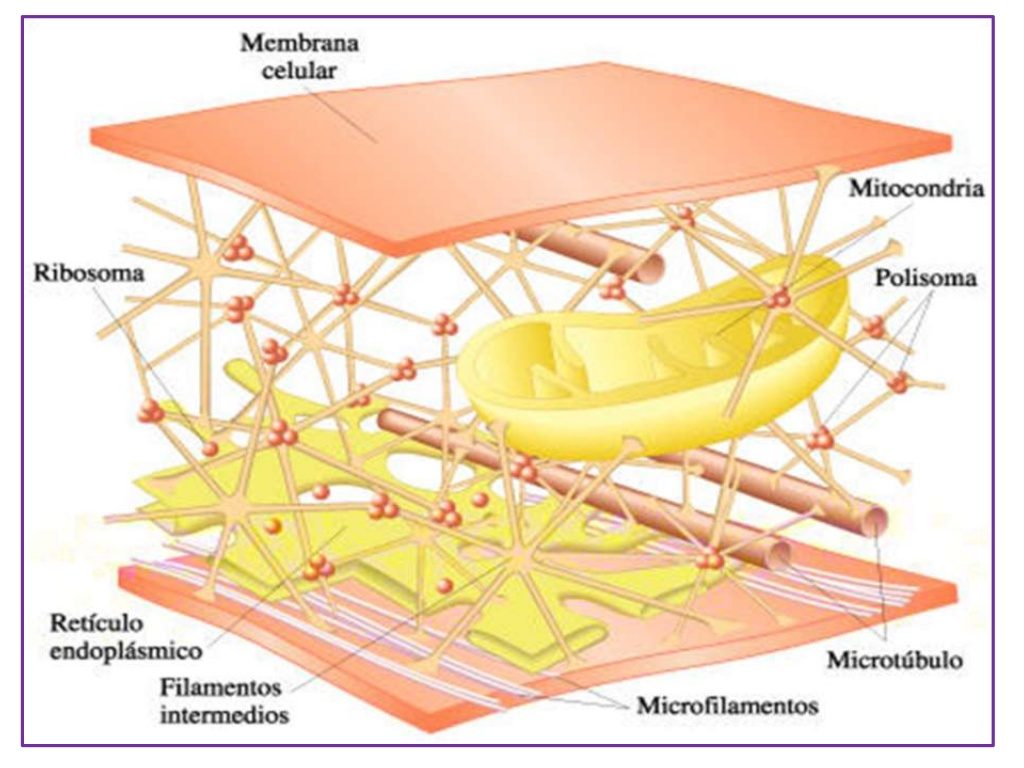


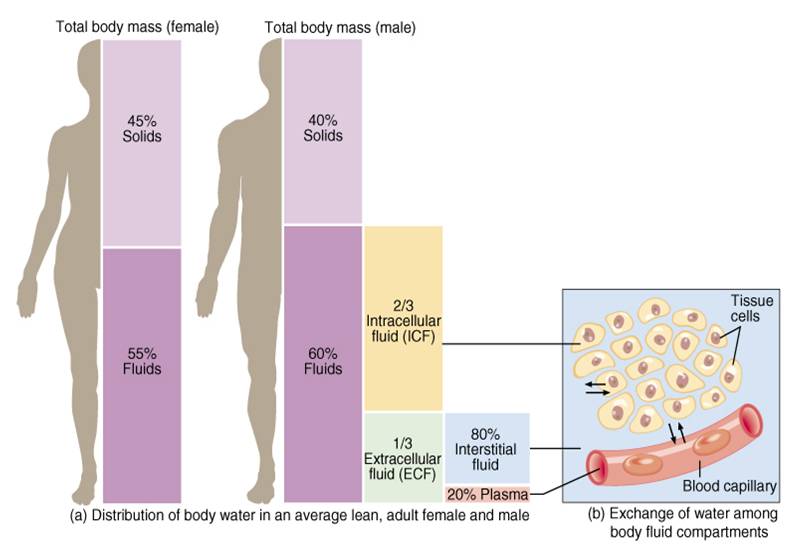

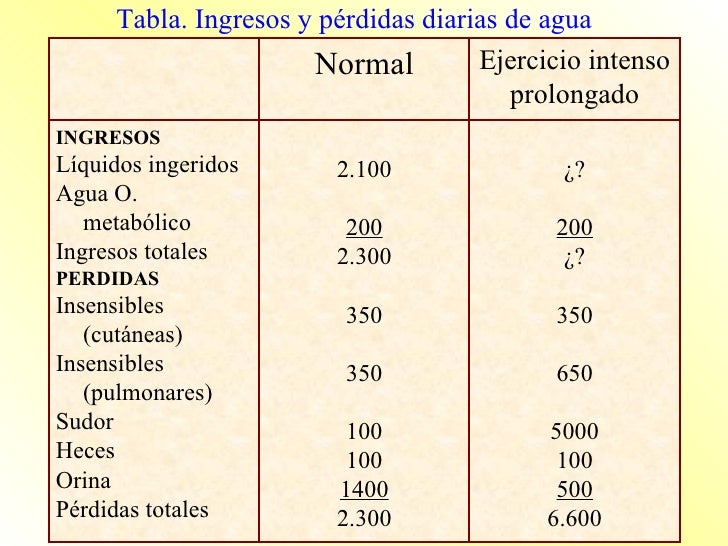

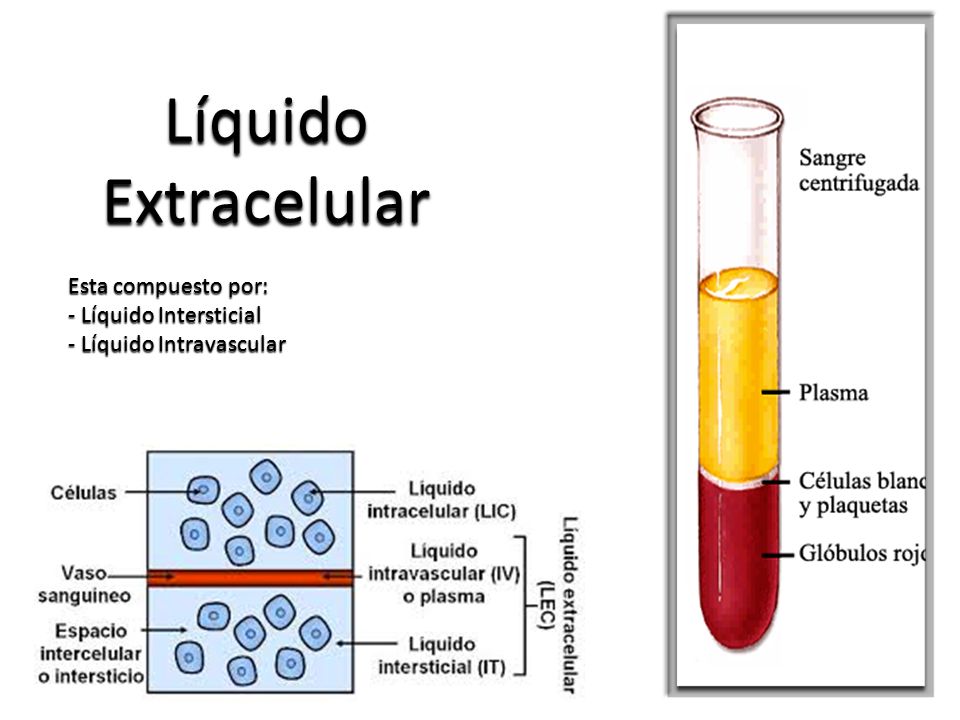


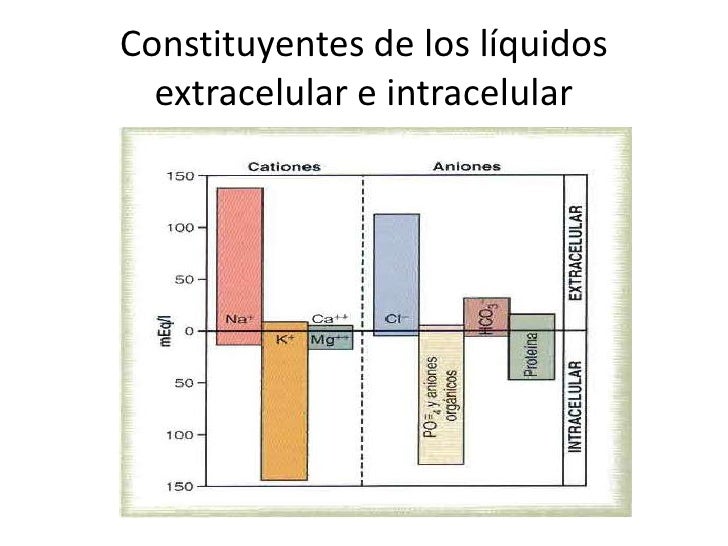
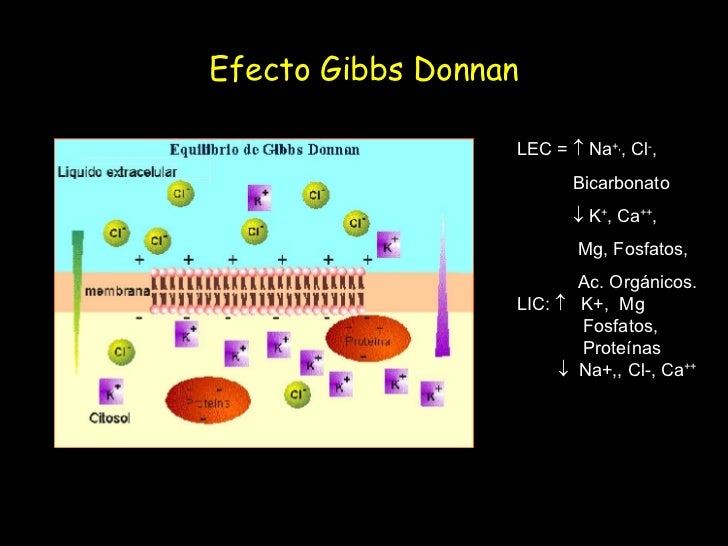
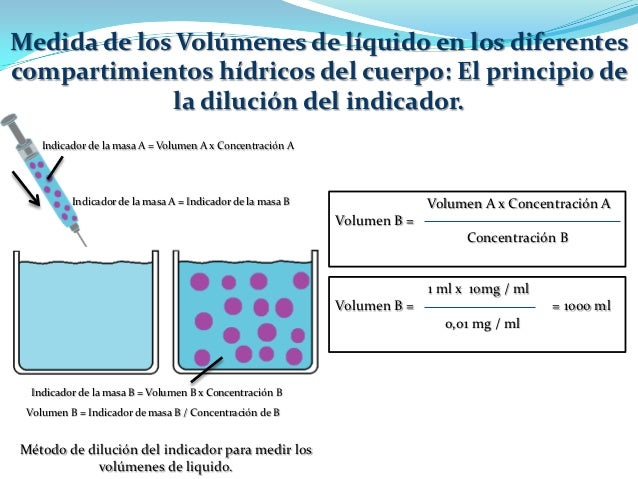



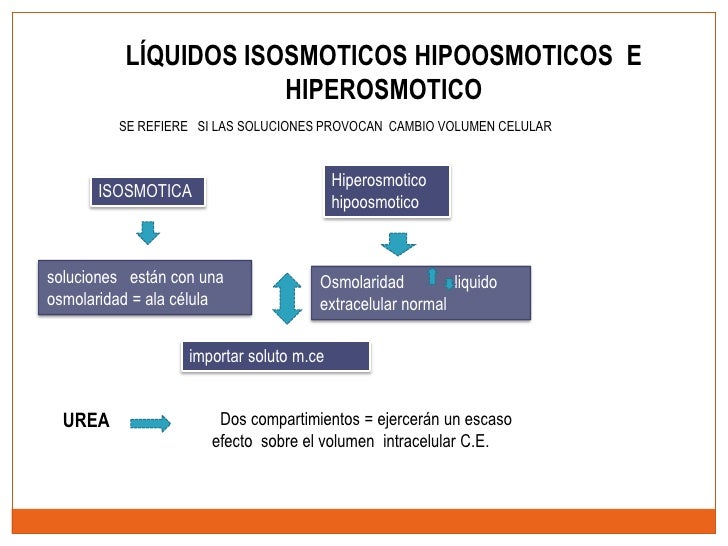






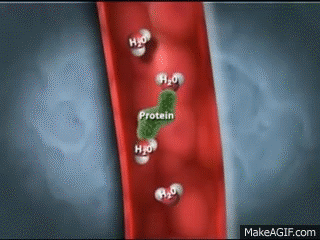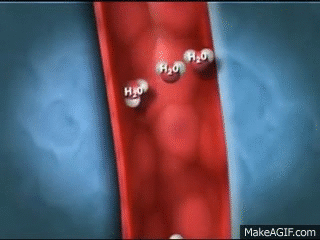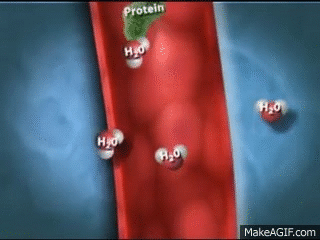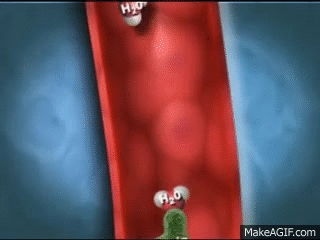



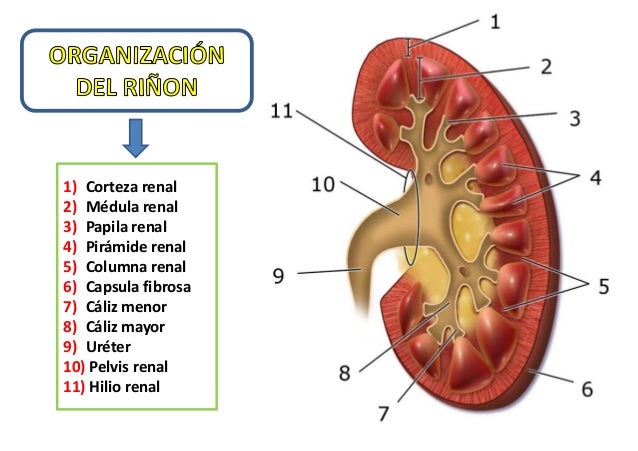


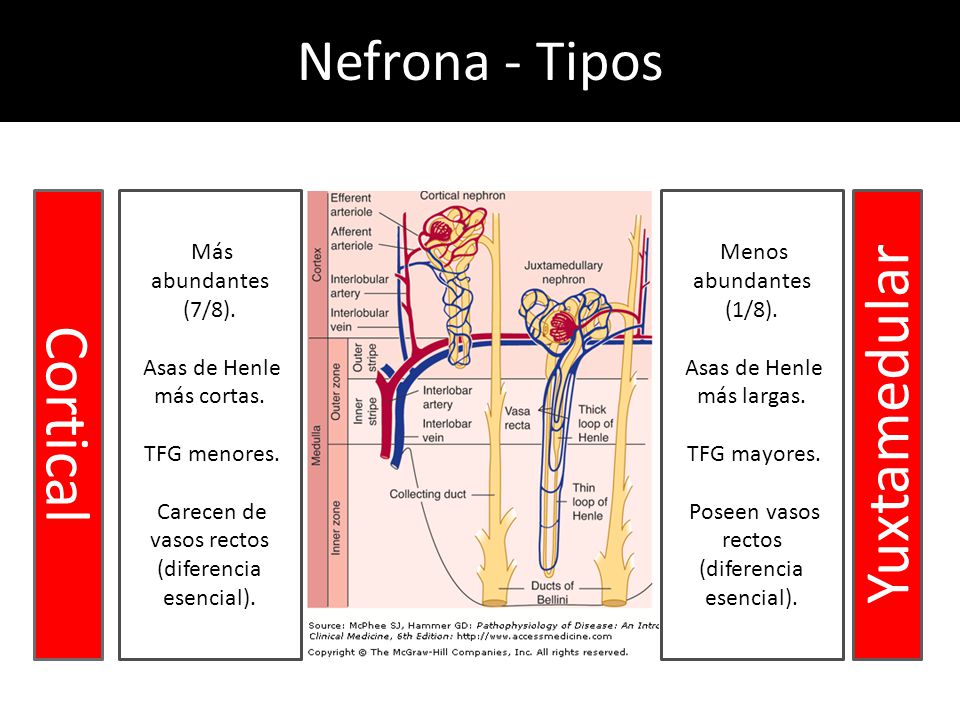




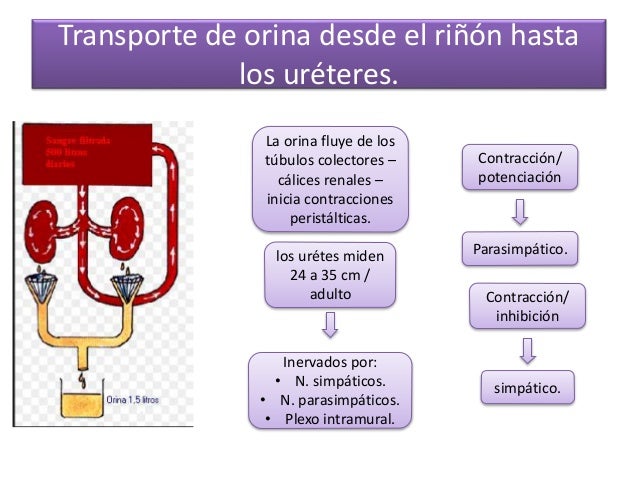
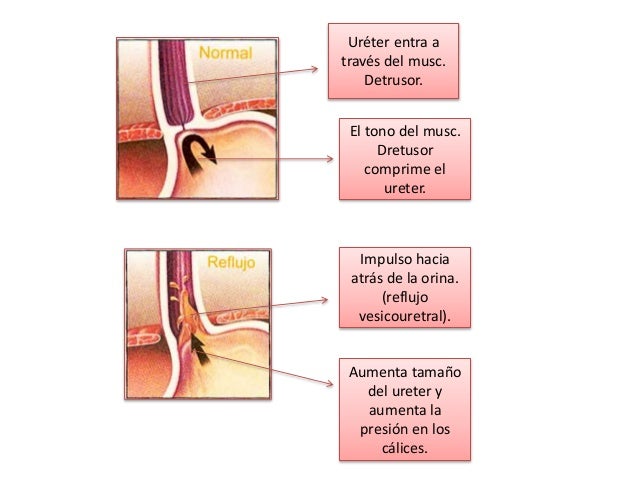

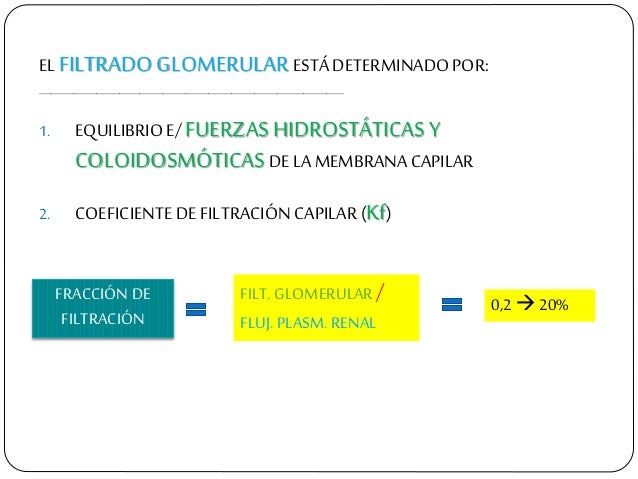
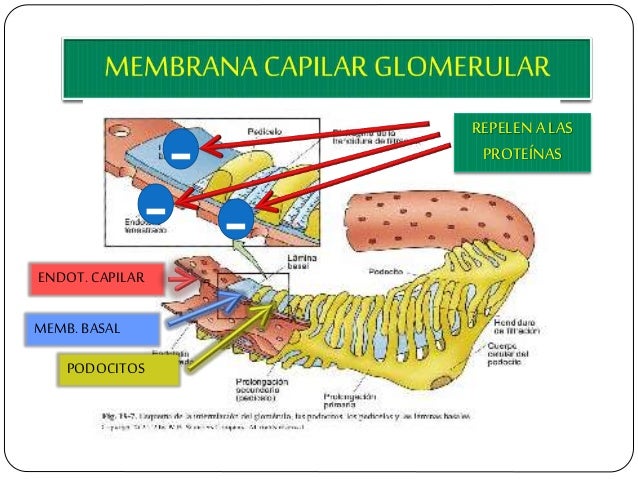
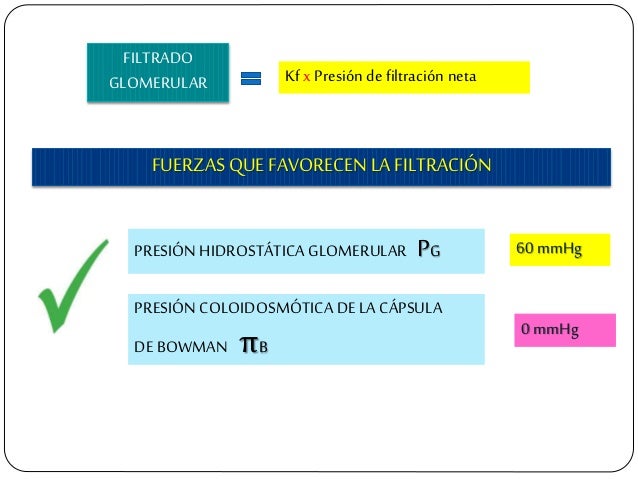
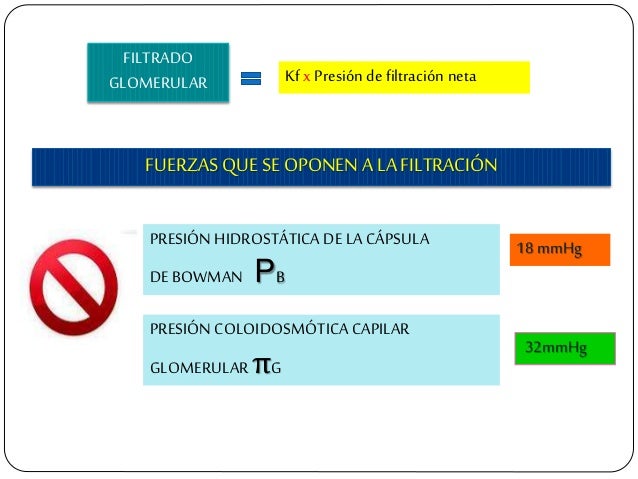
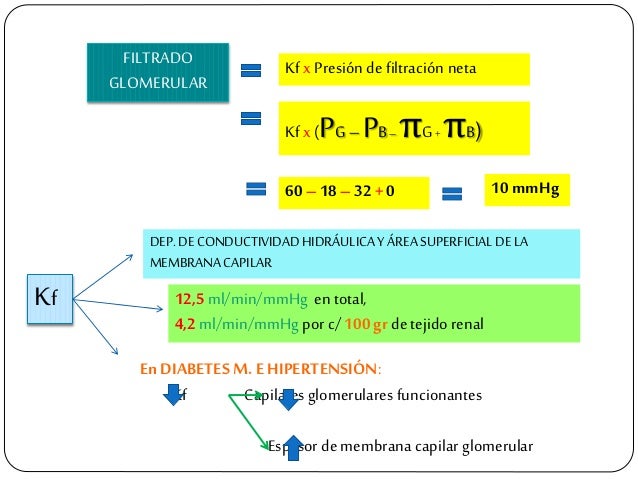
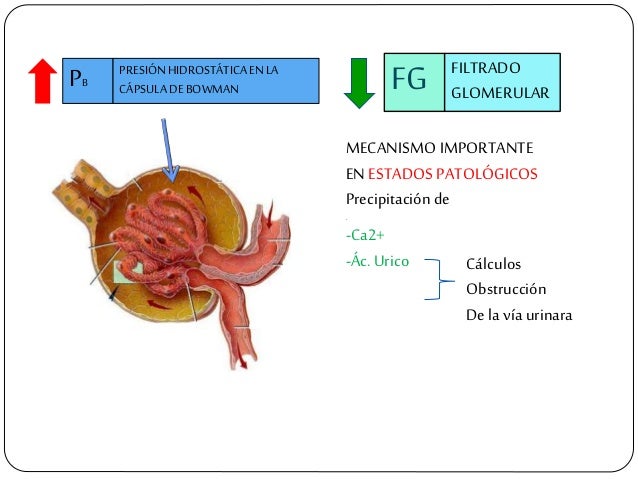
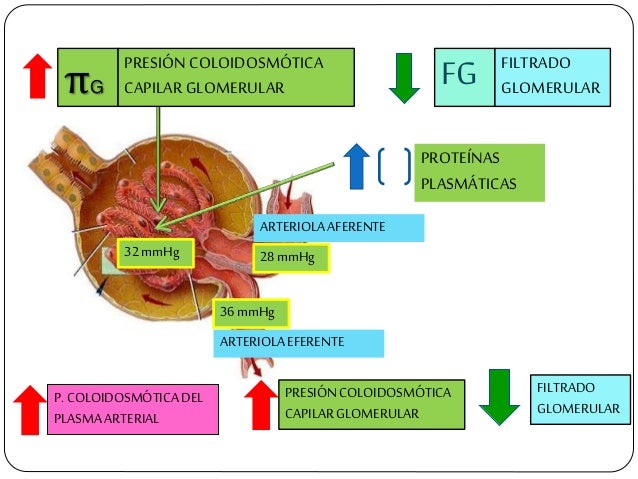
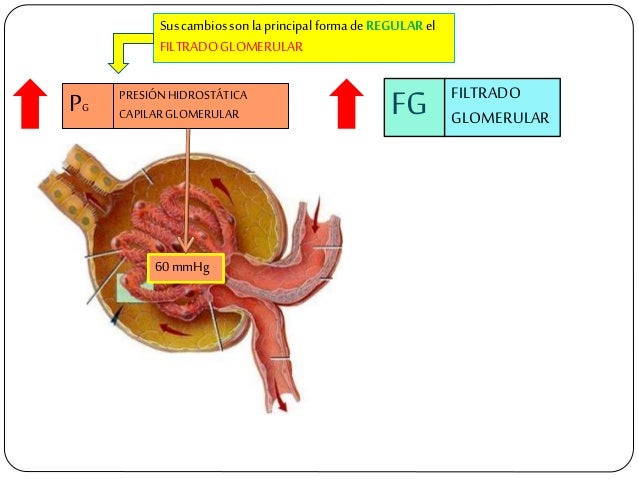
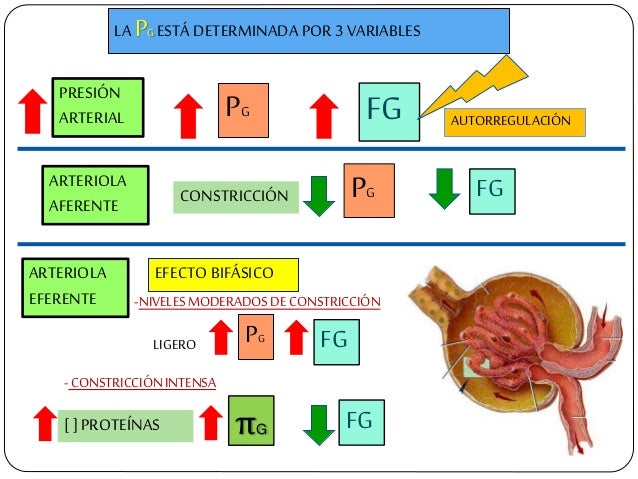
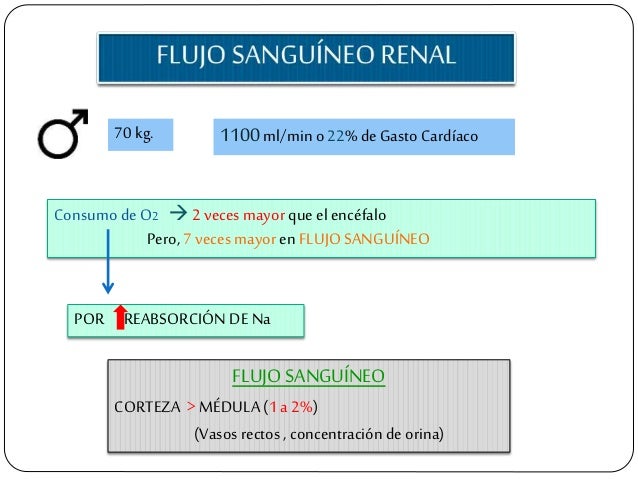
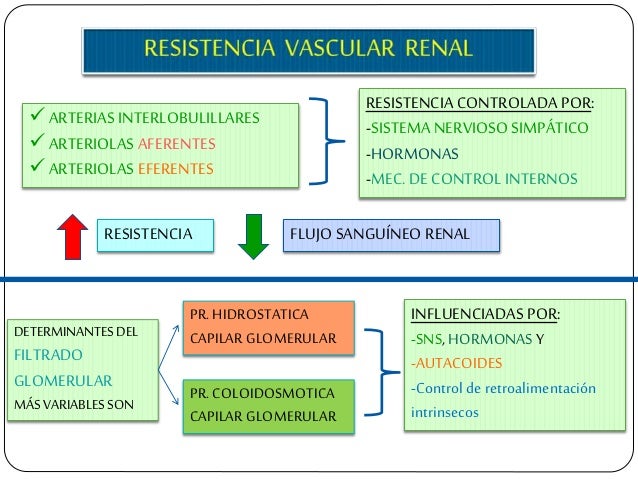
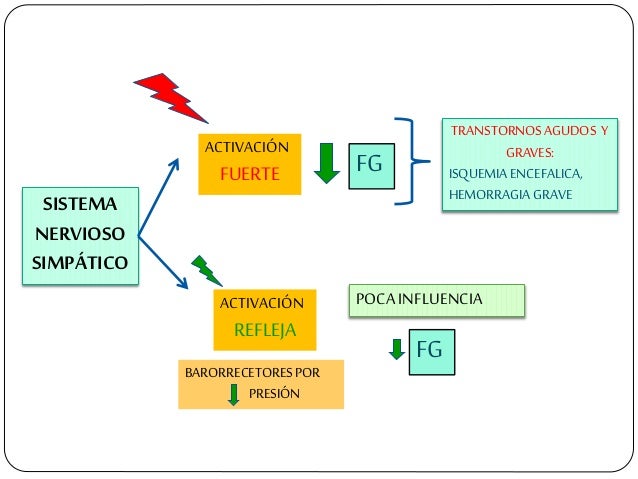
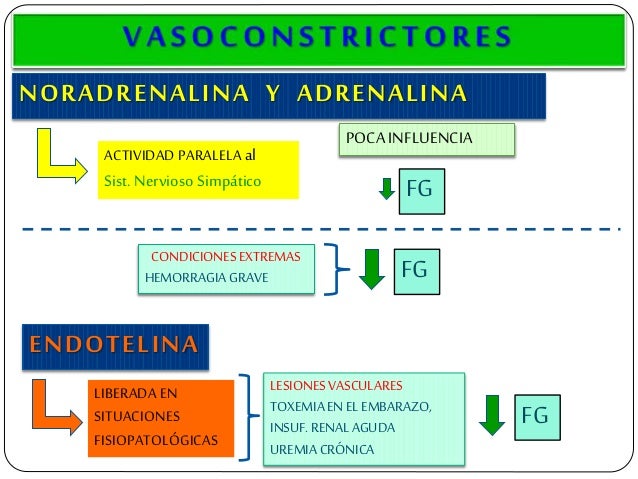
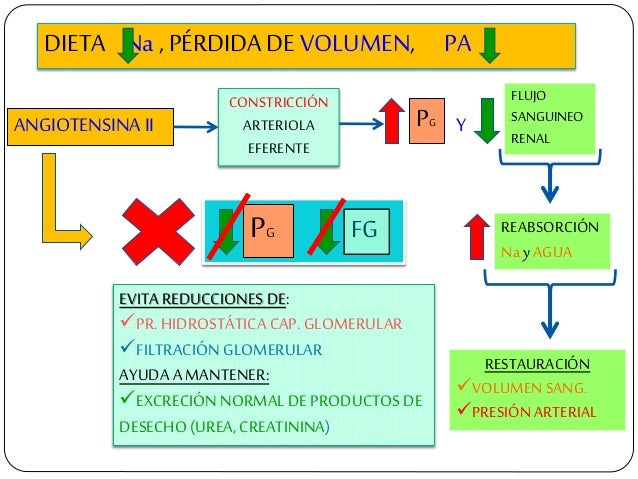
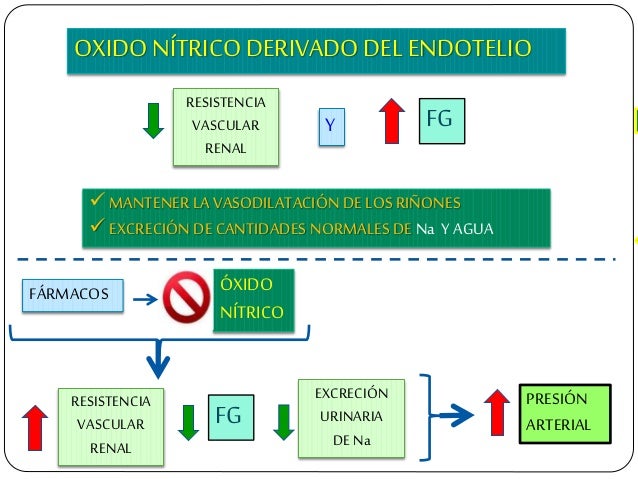
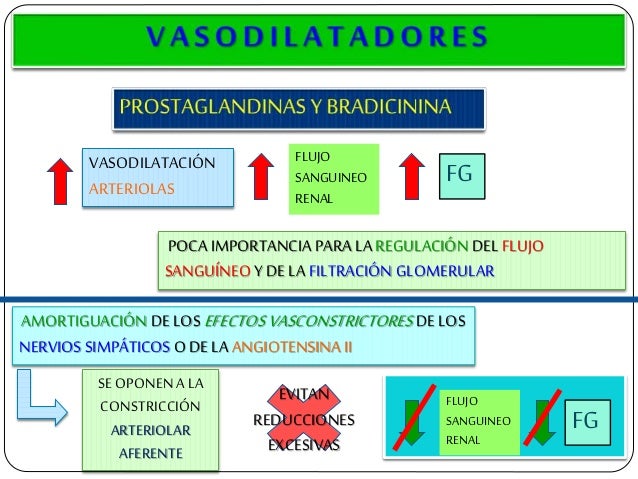
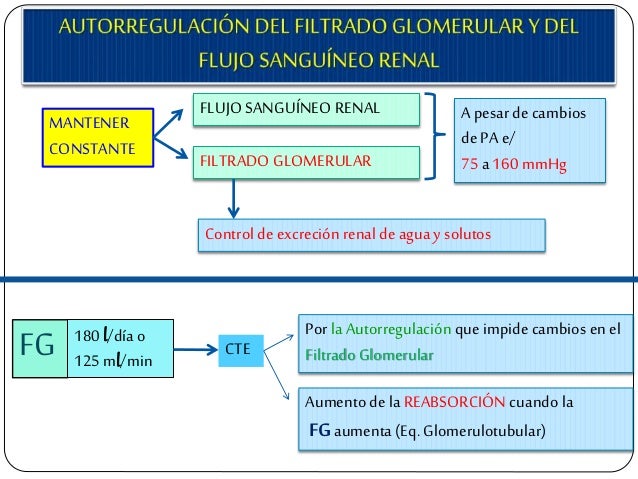
![REGULACIÃN DEL FLUJO SANGUÃNEO RENAL Y EL FILTRADO GLOMERULAR POR CAMBIOS EN LA [] deNaCl enla MÃCULA DENSA 2 Componentes ...](https://image.slidesharecdn.com/trabajo-140612075215-phpapp01/95/filtracion-glomerular-22-638.jpg?cb=1402560207)