Hay potenciales eléctricos a través de las membranas de prácticamente todas las células del cuerpo. Además, algunas células, como las células nerviosas y musculares, son capaces de generar impulsos electroquímicos rápidamente cambiantes en sus membranas, y estos impulsos se utilizan para transmitir señales a través de las membranas de los nervios y de los músculos.
En otros tipos de células, como las células glandulares, los macrófagos y las células ciliadas, los cambios locales de los potenciales de membrana también activan muchas de las funciones de las células. Este análisis se refiere a los potenciales de membrana que se generan tanto en reposo como durante la acción en las células nerviosas y musculares.

Física básica de los potenciales de membrana
Potenciales de membrana provocados por difusión
«Potencial de difusión»
producido por una diferencia de concentración iónica a los dos lados de la membrana. Debido al gran gradiente de concentración de potasio desde el interior hacia el exterior hay una intensa tendencia a que cantidades adicionales de iones potasio difundan hacia fuera a través de la membrana.
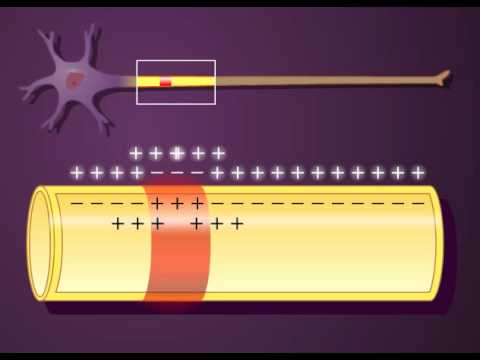
En un plazo de aproximadamente 1 ms la diferencia de potencial entre el interior y el exterior, denominada potencial de difusión, se hace lo suficientemente grande como para bloquear la difusión adicional neta de potasio hacia el exterior, a pesar del elevado gradiente de concentración iónica de potasio. En la fibra nerviosa normal del mamífero la diferencia de potencial necesaria es de aproximadamente 94 mV, con negatividad en el interior de la membrana de la fibra.
La difusión de los iones sodio de carga positiva hacia el interior crea un potencial de membrana de polaridad opuesta, con negatividad en el exterior y positividad en el interior. Una vez más el potencial de membrana se hace lo suficientemente elevado en un plazo de milisegundos como para bloquear la ulterior difusión neta de iones sodio hacia el interior; sin embargo, esta vez, en la fibra nerviosa del mamífero, el potencial es de aproximadamente 61 mV positivos en el interior de la fibra.
Relación del potencial de difusión con la diferencia de concentración: potencial de Nernst.
El nivel del potencial de difusión a través de una membrana que se opone exactamente a la difusión neta de un ion particular a través de la membrana se denomina potencial de Nernst.
La magnitud de este potencial de Nernst viene determinada por el cociente de las concentraciones de ese ion específico en los dos lados de la membrana. Cuanto mayor es este cociente, mayor es la tendencia del ion a difundir en una dirección y, por tanto, mayor será el potencial de Nernst necesario para impedir la difusión neta adicional.
Se calcula que el potencial de Nernst es de –61 mV en el interior de la membrana.

Medición del potencial de membrana
El método para medir el potencial de membrana es simple en teoría, aunque con frecuencia es difícil en la práctica debido al pequeño tamaño de la mayor parte de las fibras. La pipeta se inserta en la membrana celular hasta el interior de la fibra. Después se coloca otro electrodo, denominado «electrodo indiferente», en el líquido extracelular, y se mide la diferencia de potencial entre el interior y exterior de la fibra utilizando un voltímetro adecuado.
Se debe transferir un número increíblemente pequeño de iones a través de la membrana para establecer el «potencial en reposo» normal de –90mV en el interior de la fibra nerviosa; esto significa que sólo se debe transferir entre 1/3.000.000 a 1/100.000.000 del número total de cargas positivas del interior de la fibra. Además, un número igual de pequeño de iones positivos que se mueven desde el exterior hacia el interior de la fibra puede invertir el potencial desde –90mV hasta tanto como +35mV en tan sólo 1/10.000 de segundo.
Potencial de membrana en reposo de los nervios
El potencial de membrana en reposo de las fibras nerviosas grandes cuando no transmiten señales nerviosas es de aproximadamente –90mV. Es decir, el potencial en el interior de la fibra es 90mV más negativo que el potencial del líquido extracelular que está en el exterior de la misma.

Transporte activo de los iones sodio y potasio a través de la membrana: la bomba sodio-potasio (Na+-K+).
Se trata de una bomba electrógena porque se bombean más cargas positivas hacia el exterior que hacia el interior (tres iones Na+ hacia el exterior por cada dos iones K+ hacia el interior), dejando un déficit neto de iones positivos en el interior; esto genera un potencial negativo en el interior de la membrana celular.
La bomba Na+-K+ también genera grandes gradientes de concentración para el sodio y el potasio a través de la membrana nerviosa en reposo. Estos gradientes son los siguientes:
Na+ (exterior): 142 mEq/l Na+ (interior): 14 mEq/l
K+ (exterior): 4 mEq/l K+ (interior): 140 mEq/l
Fuga de potasio y de sodio a través de la membrana nerviosa.
«dominio de poros en tándem», canal de potasio o canal de «fuga» de potasio (K+), en la membrana nerviosa a través de la que pueden escapar iones potasio incluso en una célula en reposo.

Origen del potencial de membrana en reposo normal
Contribución de la difusión de sodio a través de la membrana nerviosa.
Esto se puede responder utilizando la ecuación de Goldman que se ha descrito previamente. Intuitivamente se puede ver que, si la membrana es muy permeable al potasio pero sólo ligeramente permeable al sodio, es lógico que la difusión del potasio contribuya mucho más al potencial de membrana que la difusión del sodio. En la fibra nerviosa normal la permeabilidad de la membrana al potasio es aproximadamente 100 veces mayor que la permeabilidad al sodio. Utilizando este valor en la ecuación de Goldman se obtiene un potencial en el interior de la membrana de –86mV,
Contribución de la bomba Na+-K+.
El hecho de que se bombeen más iones sodio hacia el exterior que iones potasio hacia el interior da lugar a una pérdida continua de cargas positivas desde el interior de la membrana; esto genera un grado adicional de negatividad (aproximadamente –4mV más) en el interior además del que se puede explicar por la difusión de manera aislada. Por tanto, el potencial de membrana neto cuando actúan todos estos mecanismos a la vez es de aproximadamente –90 mV.
En resumen, los potenciales de difusión aislados que pro- duce la difusión del sodio y del potasio darían un potencial de membrana de aproximadamente –86 mV, casi todo determinado por la difusión de potasio. Además, se generan –4mV adicionales al potencial de membrana por la acción continua de la bomba de Na+-K+ electrógena, generándose un poten- cial neto de membrana de –90 mV.
Potencial de acción nervioso
Las señales nerviosas se transmiten mediante potenciales de acción que son cambios rápidos del potencial de membrana que se extienden rápidamente a lo largo de la membrana de la fibra nerviosa. Cada potencial de acción comienza con un cambio súbito desde el potencial de membrana negativo en reposo normal hasta un potencial positivo y después termina con un cambio casi igual de rápido de nuevo hacia el potencial negativo. Para conducir una señal nerviosa el potencial de acción se desplaza a lo largo de la fibra nerviosa hasta que llega al extremo de la misma.

Las sucesivas fases del potencial de acción son las siguientes.
Fase de reposo. Este es el potencial de membrana en reposo antes del comienzo del potencial de acción. Se dice que la membrana está «polarizada» durante esta fase debido al potencial de membrana negativo de –90mV que está presente.
Fase de despolarización. En este momento la membrana se hace súbitamente muy permeable a los iones sodio, lo que permite que un gran número de iones sodio con carga positiva difunda hacia el interior del axón. El estado «polarizado» normal de –90mV se neutraliza inmediatamente por la entrada de iones sodio cargados positivamente, y el potencial aumenta rápidamente en dirección positiva. Esto se denomina despolarización.
Fase de repolarización. En un plazo de algunas diezmilésimas de segundo después de que la membrana se haya hecho muy permeable a los iones sodio, los canales de sodio comienzan a cerrarse y los canales de potasio se abren más de lo normal. De esta manera, la rápida difusión de los iones potasio hacia el exterior restablece el potencial de membrana en reposo negativo normal. Esto se denomina repolarización de la membrana.
Canales de sodio y potasio activados por el voltaje
El actor necesario en la producción tanto de la despolarización como de la repolarización de la membrana nerviosa durante el potencial de acción es el canal de sodio activado por el voltaje. Un canal de potasio activado por el voltaje también tiene una función importante en el aumento de la rapidez de la repolarización de la membrana. Estos dos cana- les activados por el voltaje tienen una función adicional a la de la bomba Na+-K+ y de los canales de fuga K+.
Canal de sodio activado por el voltaje: activación e inactivación del canal
Este canal tiene dos compuertas, una cerca del exterior del canal, denominada compuerta de activación, y otra cerca del interior, denominada compuerta de inactivación. La parte superior izquierda de la figura representa el estado de estas dos compuertas en la membrana en reposo normal, cuando el potencial de membrana es de –90 mV. En este estado la compuerta de activación está cerrada, lo que impide la entrada de iones sodio hacia el interior de la fibra a través de estos canales de sodio.
Activación del canal de sodio. Cuando el potencial de membrana se hace menos negativo que durante el estado de reposo, aumentando desde –90 mV hacia cero, finalmente alcanza un voltaje (habitualmente algún punto entre –70 y –50mV) que produce un cambio conformacional súbito en la activación de la compuerta, que bascula totalmente hasta la posición de abierta. Esto se denomina estado activado; durante este estado los iones sodio pueden atravesar el canal, aumentando la permeabilidad de la membrana al sodio hasta 500 a 5.000 veces.
Inactivación del canal de sodio. El mismo aumento de voltaje que abre la compuerta de activación también cierra la compuerta de inactivación. Sin embargo, la compuerta de inactivación se cierra algunas diezmilésimas de segundo después de que se abra la compuerta de activación. Es decir, el cambio conformacional que hace bascular la compuerta de inactivación hacia el estado cerrado es un proceso algo más lento que el cambio conformacional que abre la compuerta de activación.
Canal de potasio activado por el voltaje y su activación
Durante el estado de reposo la compuerta del canal de potasio está cerrada, lo que impide que los iones potasio pasen a través de este canal hacia el exterior. Cuando el potencial de membrana aumenta desde –90mV hacia cero, este voltaje produce una apertura conformacional de la compuerta y permite el aumento de la difusión de potasio hacia fuera a través del canal. Sin embargo, debido a la ligera demora de la apertura de los canales de potasio, en su mayor parte, se abren al mismo tiempo que están comenzando a cerrarse los canales de sodio debido a su inactivación. Por tanto, la disminución de la entrada de sodio hacia la célula y el aumento simultáneo de la salida de potasio desde la célula se combinan para acelerar el proceso de repolarización, lo que da lugar a la recuperación completa del potencial de membrana en reposo en otras pocas diezmilésimas de segundo.

Funciones de otros iones durante el potencial de acción
Hasta ahora hemos considerado sólo la función de los iones sodio y potasio en la generación del potencial de acción. Se deben considerar al menos otros dos tipos de iones: los aniones negativos y los iones calcio.
Iones con carga negativa (aniones) no difusibles en el interior del axón nervioso. En el interior del axón hay muchos iones de carga negativa que no pueden atravesar los canales de la membrana. Incluyen los aniones de las moléculas proteicas y de muchos compuestos de fosfato orgánicos, compuestos de sulfato y similares. Por tanto, estos iones negativos no difusibles son responsables de la carga negativa en el interior de la fibra cuando hay un déficit neto de iones potasio de carga positiva y de otros iones positivos.
Iones calcio. Las membranas de casi todas las células del cuerpo tienen una bomba de calcio similar a la bomba de sodio, y el calcio coopera con el sodio (o actúa en su lugar) en algunas células para producir la mayor parte del potencial de acción. Esto deja una concentración celular interna de iones calcio de aproximadamente 10–7 molar, en comparación con una concentración externa de aproximadamente 10–3 molar.
El probable mecanismo mediante el cual los iones calcio afectan a los canales de sodio es el siguiente: estos iones parecen unirse a la superficie externa de la molécula de la proteína del canal de sodio. Las cargas positivas de estos iones calcio, a su vez, alteran el estado eléctrico de la propia proteína del canal de sodio, lo que modifica el nivel de voltaje necesario para abrir la compuerta de sodio.
Inicio del potencial de acción
Un ciclo de retroalimentación positiva abre los canales de sodio.
Primero, siempre que no haya alteraciones de la membrana de la fibra nerviosa, no se produce ningún potencial de acción en el nervio normal. Sin embargo, si algún episodio produce una elevación suficiente del potencial de membrana desde –90 mV hacia el nivel cero, el propio aumento del voltaje hace que empiecen a abrirse muchos canales de sodio activados por el voltaje.
Esto per- mite la entrada rápida de iones sodio, lo que produce una elevación adicional del potencial de membrana y abre aún más canales de sodio activados por el voltaje y permite que se produzca una mayor entrada de iones sodio hacia el interior de la fibra.
Umbral para el inicio del potencial de acción.
No se producirá un potencial de acción hasta que el aumento inicial del potencial de membrana sea lo suficientemente grande como para dar origen al ciclo de retroalimentación positiva que se ha descrito en el párrafo anterior. Esto se produce cuando el número de iones Na+ que entran en la fibra supera al número de iones K+ que salen de la misma. Habitualmente es necesario un aumento súbito del potencial de membrana de 15 a 30 mV. Por tanto, un aumento súbito del potencial de membrana en una fibra nerviosa grande desde –90mV hasta aproximadamente –65mV habitualmente da lugar a la aparición explosiva de un potencial de acción. Se dice que este nivel de –65 mV es el umbral para la estimulación.

Propagación del potencial de acción
En los párrafos anteriores hemos analizado el potencial de acción que se produce en un punto de la membrana. Sin embargo, un potencial de acción que se desencadena en cualquier punto de una membrana excitable habitualmente excita porciones adyacentes de la membrana, dando lugar a la propagación del potencial de acción a lo largo de la membrana. De esta manera el proceso de despolarización viaja a lo largo de toda la longitud de la fibra. Esta transmisión del proceso de despolarización a lo largo de una fibra nerviosa muscular se denomina impulso nervioso o muscular.
Principio del todo o nada.
Una vez que se ha originado un potencial de acción en cualquier punto de la membrana de una fibra normal, el proceso de despolarización viaja por toda la membrana si las condiciones son las adecuadas, o no viaja en absoluto si no lo son. Esto se denomina principio del todo o nada y se aplica a todos los tejidos excitables normales.
Restablecimiento de los gradientes iónicos de sodio y potasio tras completarse los potenciales de acción: la importancia del metabolismo de la energía
La propagación de cada potencial de acción a lo largo de una fibra nerviosa reduce ligeramente las diferencias de concentración de sodio y de potasio en el interior y en el exterior de la membrana, porque los iones sodio difunden hacia el interior durante la despolarización y los iones potasio difunden hacia el exterior durante la repolarización. Para un único potencial de acción este efecto es tan pequeño que no se puede medir. Aun así, con el tiempo se hace necesario restablecer las diferencias de las concentraciones de membrana de sodio y de potasio. Esto se consigue por la acción de la bomba Na+-K+
Una característica especial de la bomba Na+-K+-ATPasa es que su grado de actividad se estimula mucho cuando se acumula un exceso de iones sodio en el interior de la membrana celular. De hecho, la actividad de bombeo aumenta aproximadamente en proporción a la tercera potencia de esta concentración intracelular de sodio.
Meseta en algunos potenciales de acción
En algunos casos la membrana excitada no se repolariza inmediatamente después de la despolarización; por el contrario, el potencial permanece en una meseta cerca del máximo del potencial de espiga durante muchos milisegundos, y sólo después comienza la repolarización.
La causa de la meseta es una combinación de varios factores. En primer lugar, en el proceso de despolarización del músculo cardíaco participan dos tipos de canales: 1) los canales de sodio habituales activados por el voltaje, denominados canales rápidos, y 2) los canales de calcio-sodio activados por el voltaje, que tienen una apertura lenta y que, por tanto, se denominan canales lentos.
Ritmicidad de algunos tejidos excitables: descarga repetitiva
Las descargas repetitivas autoinducidas aparecen normalmente en el corazón, en la mayor parte del músculo liso y en muchas neuronas del sistema nervioso central. Estas descargas rítmicas producen: 1) el latido rítmico del corazón; 2) el peristaltismo rítmico de los intestinos, y 3) fenómenos neuronales, como el control rítmico de la respiración.
Proceso de reexcitación necesario para la ritmicidad espontánea.
Para que se produzca ritmicidad espontánea la membrana, incluso en su estado natural, debe ser lo suficientemente permeable a los iones sodio (o a los iones calcio y sodio a través de los canales lentos de calcio- sodio) como para permitir la despolarización automática de la membrana. Después de otra demora de milisegundos o segundos la excitabilidad espontánea produce una nueva despolarización y se produce espontáneamente un nuevo potencial de acción. Este ciclo continúa de manera indefinida y produce la excitación rítmica autoinducida del tejido excitable.
El flujo aumentado de salida de iones potasio desplaza grandes cantidades de cargas positivas hacia el exterior de la membrana, dejando en el interior de la fibra una negatividad mucho mayor de lo que se produciría de otra manera. Esto continúa durante aproximadamente un segundo después de que haya finalizado el potencial de acción anterior, acercando de esta manera el potencial de membrana al potencial de Nernst del potasio. Este es un estado denominado hiperpolarización
Características especiales de la transmisión de señales en los troncos nerviosos
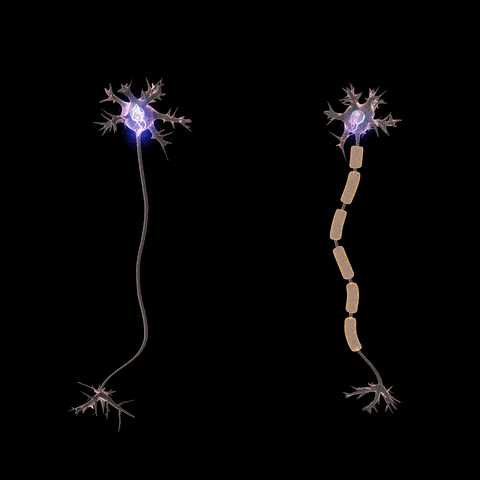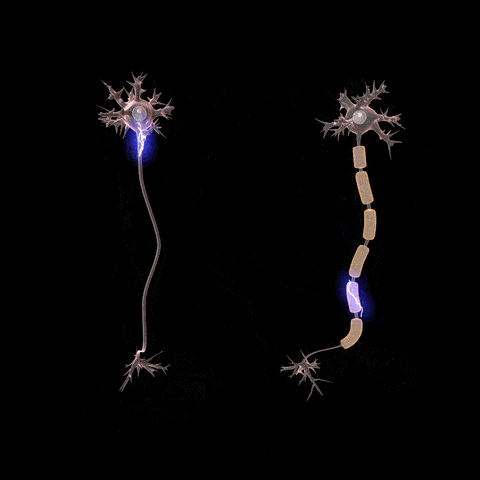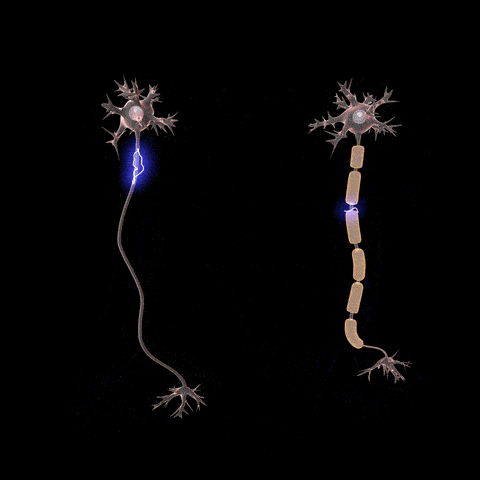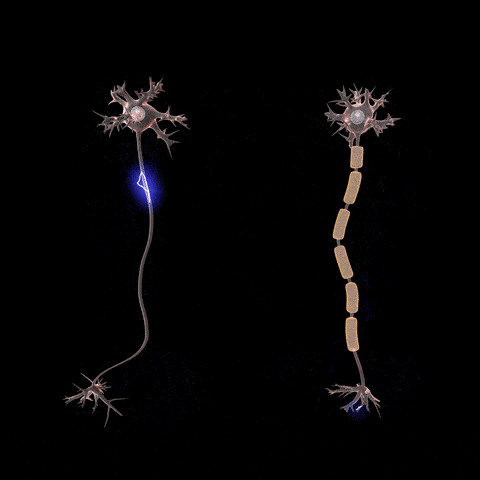
Fibras nerviosas mielinizadas y no mielinizadas.
. Las fibras grandes son mielinizadas y las pequeñas no mielinizadas. Un tronco nervioso medio contiene aproximadamente el doble de fibras no mielinizadas que mielinizadas.que con frecuencia es mucho más gruesa que el propio axón. Aproximadamente una vez cada 1 a 3 mm a lo largo de la vaina de mielina hay un nódulo de Ranvier.
Las células de Schwann depositan la vaina de mielina alrededor del axón de la siguiente manera: en primer lugar, la membrana de una célula de Schwann rodea el axón. Después, la célula de Schwann rota muchas veces alrededor del axón, depositando múltiples capas de membrana de la célula de Schwann que contiene la sustancia lipídica esfingomielina. Esta sustancia es un excelente aislante eléctrico que disminuye el flujo iónico a través de la membrana aproximadamente 5.000 veces. En la unión entre dos células de Schwann sucesivas a lo largo del axón permanece una pequeña zona no aislada de sólo 2 a 3m de longitud en la que los iones pueden seguir fluyendo con facilidad a través de la membrana del axón entre el líquido extracelular y el líquido intracelular del interior del axón. Esta zona se denomina nódulo de Ranvier.
Conducción «saltatoria» en las fibras mielinizadas de un nódulo a otro.
Aunque casi no pueden fluir iones a través de las gruesas vainas de mielina de los nervios mielinizados, pueden fluir fácilmente a través de los nódulos de Ranvier. Por tanto, los potenciales de acción se producen sólo en los nódulos; esto se denomina conducción saltatoria. Es decir, la corriente eléctrica fluye por el líquido extracelular circundante que está fuera de la vaina de mielina, así como por el axoplasma del interior del axón, de un nódulo a otro, excitando nódulos sucesivos uno después de otro. Así, el impulso nervioso recorre a saltos la fibra, lo que es el origen del término «saltatoria».

Velocidad de conducción en las fibras nerviosas.
La velocidad de conducción del potencial de acción en las fibras nerviosas varía desde tan sólo 0,25m/s en las fibras no mielinizadas pequeñas hasta 100 m/s (la longitud de un campo de fútbol en un segundo) en las fibras mielinizadas grandes.
Excitación: el proceso de generación del potencial de acción
Básicamente, cualquier factor que haga que los iones sodio comiencen a difundir hacia el interior a través de la membrana en un número suficiente puede desencadenar la apertura regenerativa automática de los canales de sodio. Esto se puede deber a un trastorno mecánico de la membrana, a los efectos químicos sobre la membrana o al paso de electricidad a través de la membrana.
Excitación de una fibra nerviosa por un electrodo metálico cargado negativamente.
El método habitual para excitar un nervio o un músculo en el laboratorio experimental es aplicar electricidad a la superficie del nervio del músculo mediante dos electrodos pequeños, uno de los cuales tiene carga negativa y el otro positiva. Cuando se hace esto la membrana excitable se estimula en el electrodo negativo.
Umbral de excitación y «potenciales locales agudos».
Un estímulo eléctrico negativo débil puede no ser capaz de excitar una fibra. Sin embargo, cuando aumenta el voltaje del estímulo se llega a un punto en el que se produce la excitación. Estos cambios locales de potencial se denominan potenciales locales agudos y, cuando no pueden generar un potencial de acción, se denominan potenciales subliminales agudos.

«Período refractario» tras un potencial de acción, durante el cual no se puede generar un nuevo estímulo
No se puede producir un nuevo potencial de acción en una fibra excitable mientras la membrana siga despolarizada por el potencial de acción precedente. El motivo de esto es que poco después del inicio del potencial de acción se inactivan los canales de sodio (o los canales de potasio, o ambos), y ninguna magnitud de la señal excitadora que se aplique a estos canales en este momento abrirá las compuertas de inactivación.
La única situación que permitirá que se vuelvan a abrir es que el potencial de membrana vuelva al nivel del potencial de membrana en reposo original o cerca del mismo. Entonces, en otra pequeña fracción de segundo se abren las compuertas de inactivación del canal y se puede iniciar un nuevo potencial de acción.
Inhibición de la excitabilidad: «estabilizadores» y anestésicos locales
Al contrario de los factores que aumentan la estabilidad nerviosa, otros factores, denominados factores estabilizadores de la membrana, pueden reducir la excitabilidad. Por ejemplo, una concentración elevada de calcio en el líquido extracelular reduce la permeabilidad de la membrana a los iones sodio y reduce simultáneamente la excitabilidad. Por tanto, se dice que el ion calcio es un «estabilizador».
