
Los genes controlan las funciones de la célula determinando qué sustancias se sintetizan dentro de la misma, es decir, qué estructuras, qué enzimas y qué productos químicos participan.
Cada gen, que corresponde a un ácido nucleico, el ácido desoxirribonucleico (ADN) el cual controla automáticamente la formación de otro ácido nucleico, el ácido ribonucleico (ARN), que después se dispersa por toda la célula para controlar la formación de una proteína específica.
El proceso completo, desde la transcripción del código genético en el núcleo hasta la traducción del código del ARN y la formación de proteínas en el citoplasma celular, se refiere como expresión génica.
Como hay aproximadamente 30.000 genes diferentes en cada célula, en teoría es posible formar un número muy grande de proteínas celulares diferentes. Algunas de las proteínas celulares son proteínas estructurales, que, asociadas a varios lípidos e hidratos de carbono, forman las estructuras de los distintos orgánulos intracelulares.
Aunque la mayoría de las proteínas son enzimas que catalizan las distintas reacciones químicas en las células. Las enzimas promueven todas las reacciones oxidativas que aportan energía a la célula y favorecen la síntesis de todos los productos químicos de la célula, como lípidos, glucógeno y trifosfato de adenosina (ATP).
Genes en el núcleo celular
En el núcleo celular hay un gran número de genes unidos por sus extremos, formando las moléculas de doble hélice largas de ADN que tienen un peso molecular que se mide por miles de millones
Bloques básicos de ADN.
La formación del ADN, se compone por el ácido fosfórico, el azúcar desoxirribosa y cuatro bases nitrogenadas (dos purínicas, adenina y guanina, y dos pirimidínicas, timina y citosina). El ácido fosfórico y la desoxirribosa forman las dos hebras helicoidales que sirven de soporte para la molécula de ADN, mientras que las bases nitrogenadas se apoyan entre las dos hebras y se conectan entre sí.
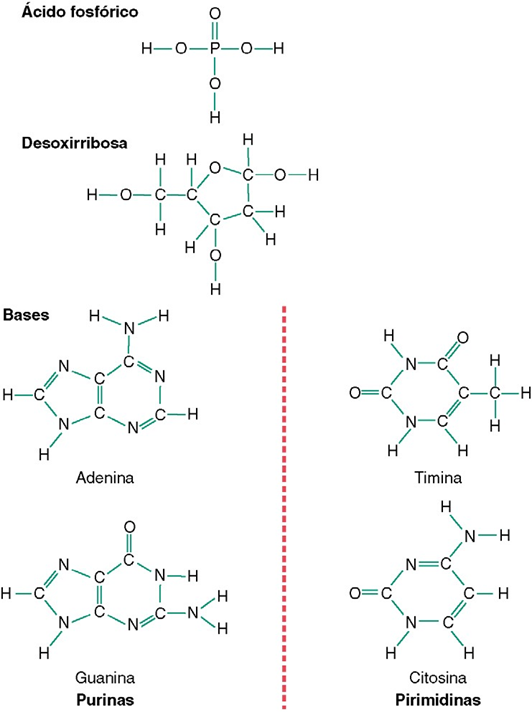
Nucleótidos.
La primera etapa en la formación del ADN consiste en combinar una molécula de ácido fosfórico, una molécula de desoxirribosa y una de las cuatro bases para formar un nucleótido ácido. De esta forma se crean cuatro nucleótidos distintos, uno para cada una de las cuatro bases, los ácidos desoxiadenílico, desoxitimidílico, desoxiguanílico y desoxicitidílico.
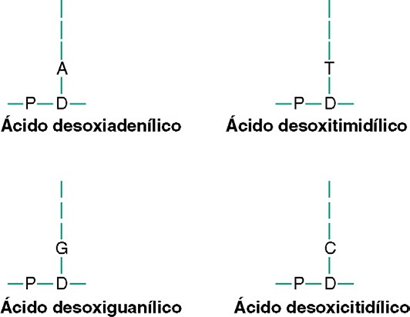
Organización de los nucleótidos para formar dos hebras de ADN
Unidas laxamente entre sí. A su vez, las dos hebras se unen laxamente entre sí mediante enlaces débiles. En el esqueleto de cada hebra de ADN está compuesto por moléculas de ácido fosfórico y desoxirribosa que se van alternando. A su vez, las bases de purina y pirimidina se unen a los lados de las moléculas de desoxirribosa. Después, las dos hebras respectivas de ADN se mantienen unidas mediante enlaces débiles de hidrógeno (líneas de puntos) entre las bases purínicas y pirimidínicas:
1. Cada base purínica de adenina de una hebra siempre se une con una base pirimidínica de timina.
2. Cada base purínica de guanina siempre se une con una base pirimidínica de citosina.
La secuencia de los pares de bases complementarios es CG, CG, GC, TA, CG, TA, GC, AT y AT. Debido a la laxitud de los enlaces de hidrógeno, las dos hebras se separan con facilidad y lo hacen muchas veces cuando realizan sus funciones en la célula.

Código genético
La importancia del ADN se debe a su capacidad para controlar la formación de las proteínas en la célula mediante un código genético. Es decir, cuando las dos hebras de la molécula de ADN se escinden quedan expuestas las bases purínicas y pirimidínicas proyectándose a un lado de cada hebra de ADN, así formándose el código genético
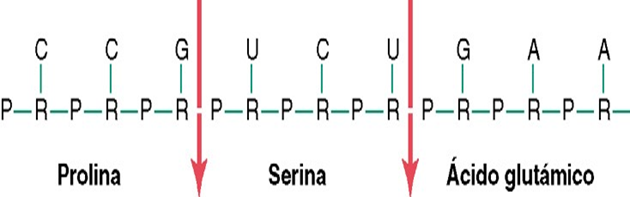
El código genético consta de «tripletes» sucesivos de bases, es decir, tres bases sucesivas componen una palabra del código. Los tripletes sucesivos controlan en último término la secuencia de aminoácidos en una molécula proteica que la célula debe sintetizar.
Un ejemplo:
El código genético GGC, AGA, CTT, estos tres tripletes respectivos son responsables de la colocación sucesiva de los tres aminoácidos, prolina, serina ácido glutámico, en una molécula de proteína de nueva formación.
El código de ADN del núcleo celular se transfiere al código de ARN en el citoplasma celular:
proceso de transcripción
Como el ADN se encuentra en el núcleo de la célula, pero la mayoría de las funciones de la célula se realizan en el citoplasma, debe haber algún mecanismo para que los genes de ADN del núcleo controlen las reacciones químicas del citoplasma, lo que se consigue mediante la intermediación de otro tipo de ácido nucleico, el ARN, cuya formación está controlada por el ADN del núcleo.
el código se transfiere al ARN en un proceso que se conoce como transcripción. A su vez, el ARN se difunde desde el núcleo a través de los poros del núcleo al compartimiento citoplasmático, donde controla la síntesis proteica.
Síntesis de ARN
Durante la síntesis de ARN las dos hebras de la molécula de ADN se separan temporalmente y una de ellas se usa como plantilla para la síntesis de una molécula de ARN.
Los tripletes del código del ADN provocan la formación de tripletes con un código complementario (o codones) en el ARN; a su vez, estos codones controlarán la secuencia de aminoácidos en una proteína que se va a sintetizar en el citoplasma celular.
Bloques básicos para la construcción del ARN.
Los bloques básicos para la construcción del ARN son prácticamente los mismos que los del ADN, excepto por dos diferencias. La primera, en la formación del ARN no se usa el azúcar desoxirribosa y en su lugar se utiliza otro azúcar que tiene una composición algo diferente, la ribosa, que contiene un ion hidroxilo extra unido a la estructura anular de la ribosa. En segundo lugar, la timina se reemplaza por otra pirimidina, uracilo.
Formación de nucleótidos de ARN.
Los bloques básicos de ADN forman los nucleótidos de ARN, exactamente igual que hemos descrito para la síntesis de ADN. En este caso, se usan también cuatro nucleótidos distintos para formar el ARN, nucleótidos que contienen las bases adenina, guanina, citosina y uracilo. Son las mismas bases que usa el ADN, excepto porque el uracilo del ARN reemplaza a la timina del ADN.
«Activación» de los nucleótidos de ARN. Es el siguiente paso de la síntesis de ARN por una enzima, la polimerasa de ARN, Se produce añadiendo a cada nucleótido dos radicales fosfato más para formar trifosfatos. Estos dos últimos fosfatos se combinan con el nucleótido mediante enlaces de fosfato de alta energía derivados del ATP celular.
El resultado de este proceso de activación es que cada uno de los nucleótidos puede disponer de grandes cantidades de energía del ATP, energía que se usa para favorecer las reacciones químicas que van añadiendo cada nuevo nucleótido de ARN al extremo de la cadena de ARN que se está desarrollando.

Montaje de la cadena de ARN a partir de los nucleótidos activados usando una cadena de ADN como plantilla:
proceso de «transcripción»
El montaje de la molécula de ARN se realiza siguiendo el proceso de una enzima, la polimerasa de ARN.
Es una gran enzima proteica que tiene muchas propiedades funcionales para la formación de la molécula de ARN, como son:
- 1) En la cadena de ADN inmediatamente contigua al gen inicial hay una secuencia de nucleótidos que se denomina promotor. La polimerasa de ARN tiene una estructura complementaria apropiada que reconoce este promotor y se une a él en un paso esencial para iniciar la formación de la molécula de ARN.
- 2) La polimerasa de ARN, después de unirse al promotor, provoca el desenrollamiento de dos vueltas de la hélice de ADN, aproximadamente, y la separación de las porciones abiertas de las dos hebras.
- 3) Después, la polimerasa se desplaza a lo largo de la hebra de ADN, desenrollando y separando temporalmente las dos hebras de ADN en cada etapa de su movimiento. A medida que se desplaza va añadiéndose en cada paso un nucleótido de ARN activado nuevo en el extremo de la cadena de ARN que se va formando, siguiendo los pasos siguientes:
*Primero, provoca la formación de un enlace de hidrógeno entre la base del extremo de la cadena de ADN y la base de un nucleótido de ARN en el nucleoplasma.
Después, la polimerasa de ARN va rompiendo dos de los tres radicales fosfato, separándolos de cada uno de estos nucleótidos de ARN y liberando grandes cantidades de energía de los enlaces de fosfato de alta energía que va rompiendo; esta energía se usa para crear el enlace covalente del fosfato que queda en el nucleótido con la ribosa en el extremo de la cadena de ARN en crecimiento.
Cuando la polimerasa de ARN alcanza el extremo del gen de ADN se encuentra con una secuencia nueva de nucleótidos de ADN que se conoce como secuencia terminadora de la cadena, que hace que la polimerasa y la cadena de ARN recién formada se separen de la cadena de ADN. Después, la polimerasa puede usarse una y otra vez para formar más cadenas de ARN.
A medida que se va formando una cadena de ARN nueva se rompen sus enlaces débiles de hidrógeno que la unen a la plantilla de ADN, porque el ADN tiene una afinidad alta para volver a unirse con su propia cadena complementaria de ADN, es decir, la cadena de ARN se separa del ADN y se libera en el nucleoplasma.

Es decir, el código presente en la cadena de ADN se transmite finalmente de forma complementariaa la cadena de ARN. Las bases de nucleótido con ribosa siempre se combinan con las bases con desoxirribosa
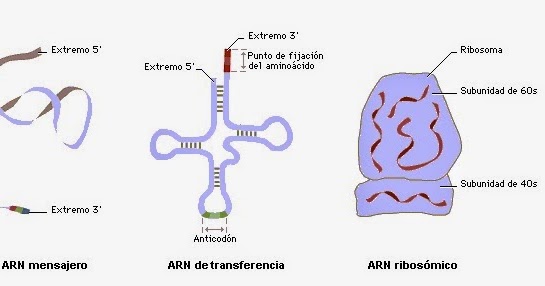
Cuatro tipos distintos de ARN.
Cada tipo de ARN tiene un papel independiente y totalmente distinto en la formación de proteínas:
1. ARN mensajero (ARNm), que transporta el código genético al citoplasma para controlar el tipo de proteína que se forma.
2. ARN de transferencia (ARNt), que transporta los aminoácidos activados a los ribosomas para usarlos en el montaje de la molécula proteica.
3. ARN ribosómico, que, junto con 75 proteínas distintas, forma ribosomas, las estructuras físicas y químicas en las que se montan realmente las moléculas proteicas.
4. MicroARN (ARNmi), que son moléculas de ARN monocatenario de 21 a 23 nucleótidos capaces de regular la transcripción y la traducción génicas.
ARN mensajero: los codones
Las moléculas de ARNm son cadenas largas y sencillas que se encuentran en suspensión en el citoplasma. Estas moléculas están compuestas por varios cientos a miles de nucleótidos de ARN en cadenas no pareadas y contienen codones que son exactamente complementarios a los tripletes del código de los genes de ADN.
Codones de ARN para los distintos aminoácidos.
| Aminoácido | Codones de ARN |
| Inicio (CI) | AUG | ||
| Parada (CT) | UAA | UAG | UGA |
Los codones de ARN de los 20 aminoácidos más frecuentes que se encuentran en las moléculas proteicas. un codón representa la señal de «iniciar la fabricación de la molécula proteica» y tres codones representan la señal de «detener la fabricación de la molécula proteica».estos dos tipos de codones se denominan CI para el «iniciador de la cadena» y CT para el «terminador de la cadena».
ARN de transferencia: los anticodones
Otro tipo de ARN que tiene un papel esencial en la síntesis proteica se conoce como ARNt, porque transfiere las moléculas de aminoácidos a las moléculas proteicas a medida que se va sintetizando la proteína.
Cada tipo de ARNt se combina específicamente con 1 de los 20 aminoácidos que se van a incorporar en las proteínas. El ARNt actúa como vehículopara transportar su tipo específico de aminoácido a los ribosomas, donde se van formando las moléculas proteicas.
En los ribosomas, cada tipo específico de ARN de transferencia reconoce un codón en particular en el ARNm y libera el aminoácido apropiado en el lugar apropiado de la cadena de la molécula proteica que se está formando.
El ARN de transferencia, que contiene sólo 80 nucleótidos, es una molécula relativamente pequeña comparada con la del ARNm. En uno de los extremos de la molécula siempre hay un ácido adenílico, en cuyo grupo hidroxilo de la ribosa del ácido adenílico se une el aminoácido transportado.
Como la función del ARNt consiste en unir un aminoácido específico a una cadena de proteína en formación, es esencial que cada tipo de ARNt también sea específico de un codón en particular del ARNm. El código específico del ARNt que le permite reconocer un codón específico es, de nuevo, un triplete de bases de nucleótidos que se denomina anticodón y se sitúa aproximadamente en la zona media de la molécula del ARNt. Durante la formación de la molécula proteica las bases del anticodón se combinan laxamente mediante enlaces hidrógeno con las bases del codón del ARNm. De esta forma, los aminoácidos respectivos se alinean uno después del otro a lo largo de la cadena del ARNm, con lo que se establece la secuencia apropiada de aminoácidos de la molécula proteica que se está formando.

ARN ribosómico
El tercer tipo de ARN de la célula es el ARN ribosómico, que constituye el 60% del ribosoma. El resto está formado por proteínas, unos 75 tipos distintos que son de tipo estructural, y también las enzimas necesarias para la fabricación de las moléculas proteicas.
El ribosoma es la estructura física del citoplasma en el que se sintetizan realmente las moléculas proteicas. No obstante, siempre funciona asociado a otros tipos de ARN: el ARNt transporta los aminoácidos al ribosoma para su incorporación en la molécula proteica en desarrollo, mientras que el ARNm proporciona la información necesaria para el secuenciado de aminoácidos en el orden adecuado para cada tipo específico de proteína que se va a fabricar.
Es decir, el ribosoma actúa como una planta de fabricación en la cual se forman las moléculas proteicas.
Formación de ribosomas en el nucléolo. Los genes de ADN que se utilizan para la formación del ARN ribosómico se sitúan en cinco pares de cromosomas del núcleo y cada uno de estos cromosomas contiene muchos duplicados de estos genes en particular, porque las funciones celulares necesitan grandes cantidades de ARN ribosómico.
A medida que se forma el ARN ribosómico se recoge en el nucléolo, una estructura especializada adyacente a los cromosomas. Cuando se están sintetizando grandes cantidades de ARN ribosómico, como sucede en las células que fabrican grandes cantidades de proteínas, el nucléolo es una estructura de gran tamaño, mientras que en las células que sintetizan pocas proteínas, el nucléolo a veces ni siquiera se ve. El ARN ribosómico se procesa especialmente en el nucléolo, donde se une a las «proteínas ribosómicas» para formar productos de condensación granulares que son las subunidades primordiales de los ribosomas.
Estas subunidades se liberan entonces desde el nucléolo y se transportan a través de los poros grandes de la cubierta nuclear hasta prácticamente todas las partes del citoplasma. Cuando estas subunidades entran en el citoplasma se reúnen para formar los ribosomas maduros y funcionales. Por tanto, las proteínas se forman en el citoplasma de la célula, pero no en el núcleo celular, porque el núcleo no contiene los ribosomas maduros.

MicroARN
Un cuarto tipo de ARN en la célula es el ARNmi. Se trata de cortos fragmentos de ARN monocatenario (de 21 a 23 nucleótidos) que regulan la expresión génica. Los ARNmi se codifican a partir del ADN transcrito de genes, pero no se traducen a proteínas y, por tanto, a menudo reciben el nombre de ARN no codificante. Los ARNmi son procesados por las células en moléculas que son complementarias al ARNm y que actúan para reducir la expresión génica. La generación de ARNmi supone un procesamiento especial de ARN precursores primarios más largos llamados ARNmi-pri, que son los transcriptos primarios del gen. Los ARNmi-pri se procesan a continuación en el núcleo celular por parte del complejo de microprocesador en pre-ARNmi, que son estructuras en horquilla de 70 nucleótidos. Estos pre-ARNmi son procesados después adicionalmente en el citoplasma por una enzima dicer específica que ayuda a ensamblar un complejo de silenciamiento inducido por ARN (RISC) y genera ARNmi.
Los ARNmi regulan la expresión génica por unión a la región complementaria del ARN y por la promoción de la represión de la traducción o degradación del ARNm antes de que pueda ser traducido por el ribosoma.
Hay ideas que los ARNmi desempeñan un papel importante en la regulación normal de la función celular, y las alteraciones en la función de los ARNmi se han asociado con enfermedades como el cáncer y las cardiopatías.

Formación de proteínas en los ribosomas: el proceso de «traducción»
Cuando una molécula de ARN mensajero entra en contacto con un ribosoma, se desplaza por el mismo a partir de un extremo predeterminado de la molécula de ARN que se especifica mediante la secuencia apropiada de las bases de ARN, el codón «iniciador de la cadena». mientras el ARN mensajero se desplaza por el ribosoma, se forma una molécula proteica, en un proceso que se conoce como traducción.
Polirribosomas. Una molécula sencilla de ARN mensajero puede formar moléculas proteicas en varios ribosomas al mismo tiempo, porque el extremo inicial de la cadena de ARN puede ir atravesando ribosomas sucesivos cuando abandona el primero. Como consecuencia de este proceso es frecuente encontrar agrupaciones de ribosomas, uniéndose entre 3 y 10 ribosomas a una única molécula de ARN mensajero al mismo tiempo. Estos grupos se conocen como polirribosomas.
Síntesis de otras sustancias en la célula Las miles de enzimas proteicas que se forman según el mismo proceso que acabamos de describir controlan esencialmente todas las reacciones químicas que tienen lugar en las células. Estas enzimas favorecen la síntesis de lípidos, glucógeno, purinas, pirimidinas y cientos de otras sustancias. Las células realizan sus múltiples funciones gracias a todas estas sustancias

Control de la función genética y actividad bioquímica de las células
Básicamente, hay dos métodos de control de las actividades químicas de la célula: 1) regulación genética, en la que se controla el grado de activación de los genes y la formación de productos génicos, y 2) regulación enzimática, en la que se controlan los niveles de actividad de las enzimas ya formadas en la célula.
Regulación genética
La regulación genética, o regulación de la expresión génica, cubre todo el proceso, desde la transcripción del código genético en el núcleo hasta la formación de proteínas en el citoplasma. La regulación de la expresión génica dota a todos los organismos vivos de la capacidad para responder a los cambios en su medio. En los animales que tienen muchos tipos diferentes de células, tejidos y órganos, la regulación diferencial de la expresión génica permite también que los numerosos tipos distintos de células del cuerpo realicen cada uno funciones especializadas. La regulación de la expresión génica puede tener lugar en cualquier punto de las rutas de transcripción, procesamiento de ARN y traducción.
Control de las funciones intracelulares mediante la regulación enzimática
Además del control de las funciones de la célula mediante la regulación genética, algunas actividades celulares están controladas por inhibidores o activadores intracelulares que actúan directamente sobre las enzimas intracelulares específicas. La regulación enzimática representa una segunda categoría de mecanismos por los cuales se pueden controlar las funciones bioquímicas celulares.
Inhibición enzimática. Algunas de las sustancias químicas formadas en la célula ejercen una retroalimentación directa inhibiendo los sistemas enzimáticos específicos que los sintetizan. Casi siempre, el producto sintetizado actúa sobre la primera enzima de una secuencia en lugar de sobre las enzimas sucesivas, uniéndose directamente a ella y provocando un cambio conformacional alostérico que la inactiva. Se puede reconocer fácilmente la importancia de la inactivación de la primera enzima, ya que impide la acumulación de los productos intermedios que no se están usando.
La inhibición enzimática es otro ejemplo de control mediante retroalimentación negativa, responsable del control de las concentraciones intracelulares de muchos aminoácidos, purinas, pirimidinas, vitaminas y otras sustancias.
Activación enzimática. Las enzimas que están normalmente inactivas se activan cuando es necesario
Se produce durante la formación de purinas y pirimidinas. La célula necesita estas sustancias en una cantidad aproximadamente igual para la formación de ADN y ARN. Las purinas formadas inhiben las enzimas que se requieren para la formación de nuevas purinas, aunque activan las enzimas necesarias para la formación de pirimidinas. Por el contrario, las pirimidinas inhiben sus propias enzimas, pero activan las enzimas de las purinas. De esta forma, hay un traspaso continuado entre los sistemas que sintetizan ambas sustancias, con lo que se producen cantidades casi iguales en todo momento.
Los genes pueden activarse o inhibirse, al igual que los sistemas enzimáticos pueden activarse o inhibirse. Estos mecanismos reguladores actúan especialmente como sistemas de control de retroalimentación que vigilan continuamente la composición bioquímica de la célula y hacen las correcciones que se van necesitando. Pero, a veces, las sustancias del exterior de la célula también controlan las reacciones bioquímicas intracelulares al activar o inhibir uno o más de los sistemas intracelulares de control.

El sistema genético de ADN también controla la reproducción celular
La reproducción celular es otro ejemplo del papel ubicuo que tiene el sistema genético del ADN en todos los procesos vitales. Los genes y sus mecanismos reguladores determinan las características de crecimiento de las células y también si se dividen para formar nuevas células y cuándo. De esta manera, el sistema genético, tan importante, controla cada etapa del desarrollo del ser humano, desde el óvulo unicelular fertilizado hasta todo un organismo funcionante. Es decir, si hay un eje central a la vida, es el sistema genético del ADN.
Ciclo vital de la célula. El ciclo vital de una célula es el período que transcurre desde el inicio de la reproducción celular hasta el inicio de la siguiente reproducción celular. Cuando las células de los mamíferos no están inhibidas, se reproducen tan rápidamente como pueden y su ciclo vital puede ser tan sólo de 10 a 30h y termina por una serie de sucesos físicos diferenciados que se denominan mitosis y que tienen como finalidad lograr la división de la célula en dos células hijas nuevas.
La mitosis en sí misma dura sólo unos 30min, por lo que más del 95% del ciclo vital de las células está representado por el intervalo entre las mitosis, o interfase, incluso en las células que se reproducen con mayor rapidez.
Excepto en determinadas condiciones especiales de reproducción celular rápida, los factores inhibidores casi siempre disminuyen la velocidad o detienen el ciclo vital no inhibido, es decir, las distintas células del organismo tienen realmente ciclos vitales que varían de tan sólo 10 h en las células de la médula ósea, sometidas a una estimulación elevada, a un ciclo vital que dura prácticamente la vida del sujeto en la mayoría de las células nerviosas.
La reproducción celular comienza con la replicación del ADN
Como sucede en la mayoría de los demás sucesos vitales importantes, la reproducción comienza en el núcleo. El primer paso consiste en la replicación (duplicación) de todo el ADN de los cromosomas y sólo después puede tener lugar la mitosis.
El ADN comienza a duplicarse entre 5 y 10h antes de la mitosis y se completa en 4-8h. El resultado neto es que se producen dos réplicas exactas de todo el ADN. Estas réplicas se convierten en el ADN de las dos células hijas nuevas que se formarán en la mitosis. Después de esta replicación hay otro periodo de 1-2 h antes de que comience bruscamente la mitosis. Durante este período comienzan los cambios preliminares que conducirán a la mitosis.

Reparación de ADN, «corrección de lectura» y«mutaciones» del ADN.
Durante la hora, más o menos, que transcurre entre la replicación del ADN y el comienzo de la mitosis hay un período de reparación muy activa y «corrección de lectura» de las cadenas de ADN; es decir, siempre que se hayan emparejado nucleótidos de ADN incorrectos con la cadena original que sirve de plantilla actúan unas enzimas especiales que cortan las zonas defectuosas y las reemplazan con los nucleótidos complementarios apropia- dos. Para ello se utilizan las mismas polimerasas del ADN y ADN ligasas que se usan en la replicación. Este proceso de reparación del ADN se conoce como corrección de lectura.
Debido a los procesos de reparación y corrección de lectura, el proceso de transcripción comete errores pocas veces pero, cuando lo hace, el error se denomina mutación. La mutación provoca la formación de alguna proteína anormal en la célula en lugar de la proteína necesaria, lo que conduce a funciones celulares anormales y, en ocasiones, incluso a la muerte celular. Aun así, y dado que hay 30.000 genes o más en el genoma humano y que el período de una generación humana a otra es de unos 30 años, tendríamos que esperar hasta 10 o más mutaciones en el paso del genoma de un padre a su hijo. No obstante, como protección el genoma humano está representado por dos conjuntos independientes de cromosomas con genes casi idénticos, por lo que el niño dispone casi siempre de un gen funcional de cada par, a pesar de las mutaciones.

Mitosis celular
El proceso real por el que la célula se divide en dos células nuevas es la mitosis. Una vez que cada cromosoma se ha replicado para formar las dos cromátidas, en muchas células la mitosis se produce automáticamente en 1 o 2 h.
Aparato mitótico: función de los centriolos. Uno de los primeros pasos de la mitosis tiene lugar en el citoplasma, al final de la interfase, en torno o en unas pequeñas estructuras denominadas centriolos, dos pares de centríolos se mantienen estrechamente unidos cerca de un polo del núcleo (estos centríolos, como el ADN y los cromosomas, también se han replicado durante la interfase, habitualmente poco después de la replicación del ADN). Cada centríolo es un pequeño organismo cilíndrico en torno a 0,4 mm de largo y 0,15 mm de diámetro y está formado principalmente por nueve estructuras tubulares paralelas dispuestas en forma de un cilindro. Los dos centríolos de cada par se disponen en ángulos rectos entre sí y cada par de centríolos, junto al material pericentriolar unido a él, compone el centrosoma.
Poco antes de que tenga lugar la mitosis, los dos pares de centríolos comienzan a separarse uno de otro, lo que se debe a la polimerización de las proteínas de los microtúbulos que crecen entre los pares respectivos de centríolos y los separan. Al mismo tiempo, crecen radialmente otros microtúbulos que alejan los pares de centríolos, formando una estrella a modo de soporte, que se conoce como áster, en cada extremo de la célula. Algunas de las puntas del áster
penetran en la membrana nuclear y permiten separar los dos conjuntos de cromátidas durante la mitosis. El complejo de microtúbulos que se extiende entre los dos nuevos pares de centríolos es el huso, y todo el conjunto de microtúbulos más los dos pares de centríolos se denomina aparato mitótico.
Profase. La primera etapa de la mitosis, denominada profase. Mientras se forma el haz, los cromosomas del núcleo (que en la interfase corresponden a hebras laxamente enrolladas) se condensan en cromosomas bien definidos.
Prometafase. las puntas de los microtúbulos en crecimiento del áster se fragmentan en la cubierta nuclear. Al mismo tiempo, los múltiples microtúbulos del áster se unen a las cromátidas en los centrómeros, donde las cromátidas pareadas aún están unidas entre sí; a continuación, los túbulos tiran de una cromátida de cada par, alejando cada una hacia el polo celular correspondiente.
Metafase. Durante la metafase los dos ásteres del aparato mitótico se separan, lo que parece suceder porque las puntas de los microtúbulos de ambos, donde se imbrican entre sí para formar el huso mitótico, realmente se empujan mutuamente. Por este motivo, parece que hay unas moléculas proteicas contráctiles diminutas, o motores moleculares, que tal vez estén formadas por la proteína muscular actina que se extienden entre las puntas respectivas para, utilizando una acción paulatina como en el músculo, deslicen activamente las puntas en dirección opuesta una de otra. Simultáneamente, los microtúbulos insertados en las cromátidas tiran fuertemente de ellas hasta el centro de la célula, alineándolas para formar el plano ecuatorial del huso mitótico.
Anafase. Durante esta fase las dos cromátidas de cada cromosoma son separadas en el centrómero. Se separan los 46 pares de cromátidas y se forman dos juegos independientes de 46 cromosomas hijos. Cada uno de ellos es empujado hacia cada uno de los ásteres de la mitosis, a medida que los dos polos respectivos de la célula en división se van separando entre sí.
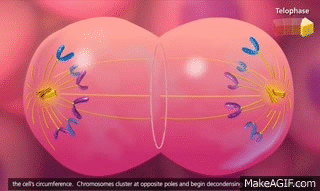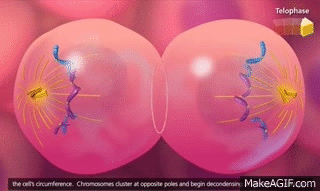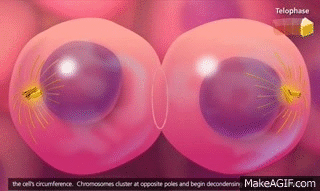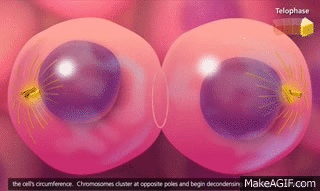
Telofase. En la telofase los dos juegos de cromosomas hijos se separan completamente. A continuación, el aparato mitótico se disuelve y se desarrolla una nueva membrana nuclear que rodea cada grupo de cromosomas. Esta membrana se forma a partir de porciones del retículo endoplásmico que ya están presentes en el citoplasma. Poco después, la célula se divide en dos, en la zona media entre los dos núcleos, como consecuencia de la formación de un anillo contráctil de microfilamentos compuestos por actina y, probablemente, miosina (las dos proteínas contráctiles del músculo) en la unión de las células nuevas que se están desarrollando, anillo que las termina separando.
Regulación del tamaño de la célula. El tamaño de la célula está determinado casi en su totalidad por la cantidad de ADN funcionante que hay en el núcleo. Si el ADN no se replica, la célula crece hasta un tamaño determinado y después lo mantiene, mientras que también es posible que, usando el quimioterápico colchicina, se impida la formación del huso mitótico y, por tanto, la mitosis aunque continúe la replicación del ADN. En este caso, el núcleo contiene una cantidad de ADN bastante mayor de lo que normalmente contendría y la célula crece hasta un tamaño proporcionalmente mayor. Se supone que este efecto es consecuencia, sencillamente, del aumento de la producción de ARN y de las proteínas celulares, lo que, a su vez, hace que el tamaño de la célula aumente más.
Diferenciación celular
Una característica especial del crecimiento y división celular es la diferenciación celular, que se refiere a los cambios de las propiedades físicas y funcionales de las células a medida que proliferan en el embrión para formar las distintas estructuras y órganos corporales.
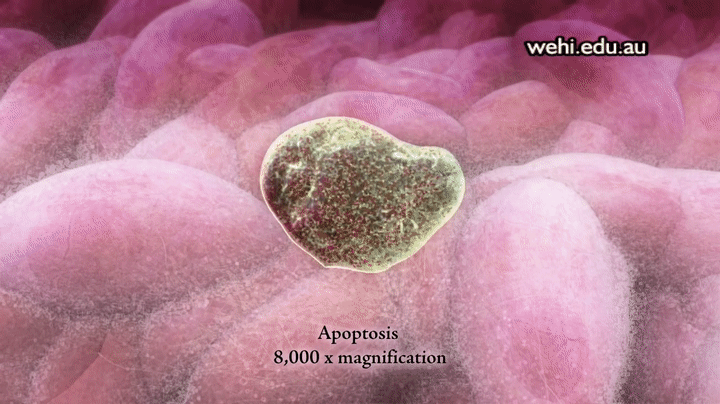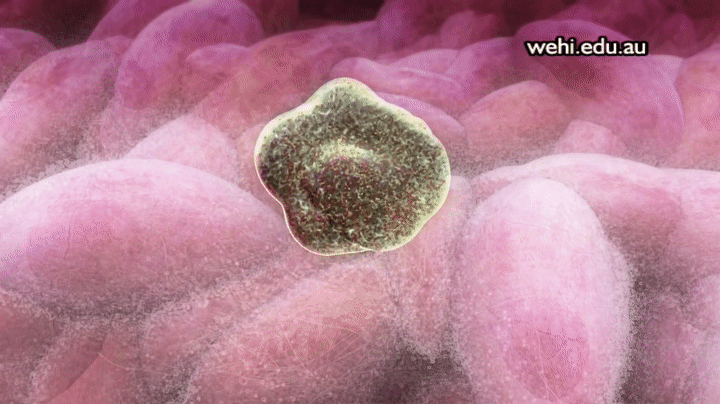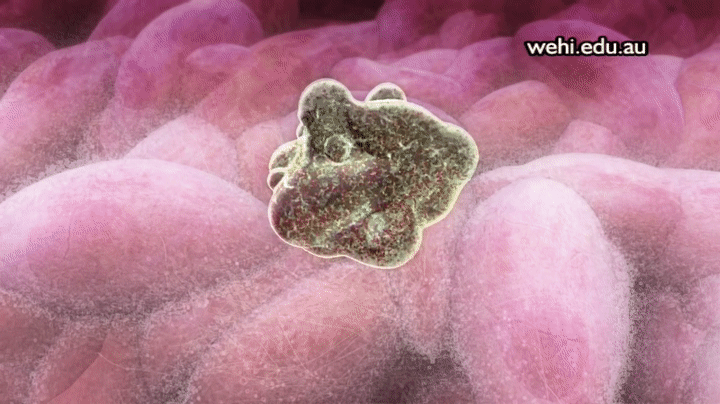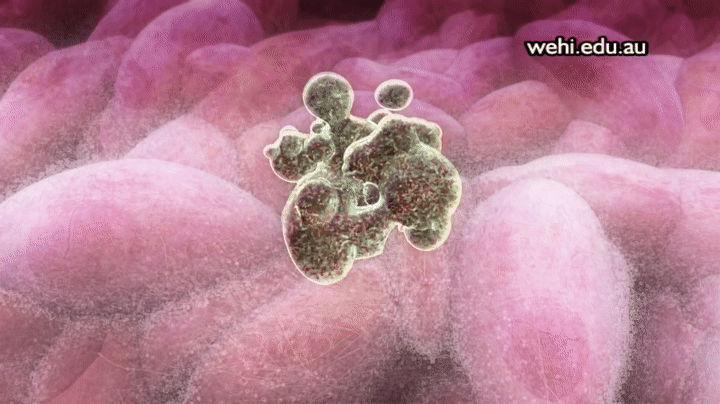
Apoptosis: muerte celular programada
Los 100 billones de células del organismo forman parte de una comunidad muy bien organizada en la que el número total de células está regulado no sólo por el control de la velocidad de división celular, sino también por el control de la velocidad de la muerte celular. Cuando las células ya no se necesitan, o cuando se convierten en una amenaza para el organismo, sufren una muerte celular programada suicida, o apoptosis. Este proceso implica una cascada proteolítica específica que hace que la célula se encoja y condense para desmontar su citoesqueleto y alterar su superficie de tal forma que una célula fagocítica cercana, como un macrófago, se puede unir a la membrana celular y digerir la célula.
Al contrario de la muerte programada, las células que mueren como consecuencia de una lesión aguda se hinchan y estallan debido a la pérdida de la integridad de la membrana celular, un proceso que se denomina necrosis celular. Las células necróticas vierten su contenido haciendo que la inflamación y la lesión se extiendan a las células vecinas. No obstante, la apoptosis es una muerte celular metódica que da lugar al desmontaje y fagocitosis de la célula antes de que se produzca ninguna fuga de su contenido, por lo que las células vecinas se mantienen sanas.
La apoptosis se inicia mediante la activación de una familia de proteasas que se conocen como caspasas. Se trata de unas enzimas que se sintetizan y almacenan en la célula en forma de procaspasas inactivas. Los mecanismos de activación de las caspasas son complejos pero, una vez activadas, las enzimas se escinden y activan otras procaspasas, activando una cascada que rompe rápidamente las proteínas del interior de la célula. De esta forma, la célula se desmantela a sí misma y sus restos se digieren rápidamente en las células fagocíticas vecinas.
Cáncer
El cáncer se debe en todos o casi todos los casos a la mutación o a alguna otra activación anormal de los genes celulares que controlan el crecimiento y la mitosis celular. Los genes anormales se denominan oncogenes y se han descubierto hasta 100 tipos distintos.
En todas las células también hay antioncogenes, que suprimen la activación de los oncogenes específicos, es decir, la pérdida o inactivación de los antioncogenes permite la activación de los oncogenes que conduce al cáncer.
La única posibilidad es que se produzca todo lo que se necesita para que tenga lugar la mutación, por lo que podemos suponer que un gran número de cánceres son, simplemente, el resultado de un suceso desafortunado.
No obstante, la probabilidad de mutaciones aumenta muchas veces cuando una persona se expone a determinados factores químicos, físicos o biológicos, como son los siguientes:
Es bien sabido que la radiación ionizante, como los rayos X, los rayos gamma y la radiación de partículas procedentes de sustancias radiactivas, e incluso la luz ultravioleta, predispone al cáncer. Los iones formados en las células tisulares bajo la influencia de este tipo de radiación son muy reactivos y pueden romper las cadenas de ADN, con lo que se provocan muchas mutaciones.
Algunas sustancias químicas también tienen una mayor propensión a provocar mutaciones. Hace tiempo se des- cubrió que hay varios derivados del colorante anilina que pueden provocar cáncer, por lo que los trabajadores de plantas químicas que producen este tipo de sustancias tienen una predisposición especial a desarrollar un cáncer si no usan protección. Las sustancias químicas que provocan la mutación se denominan carcinógenos. Los carcinógenos que actualmente provocan el mayor número de muertes son los contenidos en el humo de los cigarrillos, que provocan aproximadamente la cuarta parte de todas las muertes por cáncer.
Los irritantes físicos también provocan cáncer, como sucede durante la abrasión continuada del revestimiento del aparato digestivo por algunos alimentos. El daño de los tejidos conduce a una sustitución mitótica rápida de las células. Cuanto más rápida sea la mitosis, mayor será la probabilidad de mutación.
En muchas familias hay una importante tendencia hereditaria al cáncer, lo que es consecuencia de que la mayoría de los cánceres requieren no sólo una mutación, sino dos o más antes de que aparezca el cáncer. En esas familias particularmente predispuestas al cáncer se supone que ya han mutado uno o más genes cancerosos en el genoma heredado, por lo que en estas familias tienen que producirse muchas menos mutaciones adicionales antes de que comience a crecer un cáncer.

Características invasivas de la célula cancerosa.
Las principales diferencias entre la célula cancerosa y la célula normal son las siguientes:
- la célula cancerosa no respeta los límites habituales del crecimiento celular, ya que, presumiblemente, no requieren los mismos factores de crecimiento que son necesarios para el crecimiento de las células normales.
- Las células cancerosas son bastante menos adhesivas entre sí que las células normales, por lo que tienden a dispersarse por los tejidos, entrar al torrente sanguíneo y transportarse por el organismo, donde forman nidos para crecimientos cancerosos nuevos.
- Algunos cánceres también producen factores angiogénicos que provocan el crecimiento de muchos vasos sanguíneos nuevos dentro del cáncer, por lo que aportan los nutrientes necesarios para el crecimiento celular.

¿Por qué matan las células cancerosas?
La respuesta a esta pregunta suele ser sencilla. El tejido canceroso compite con los tejidos normales para lograr los nutrientes. Como las células cancerosas continúan proliferando indefinidamente, su número se multiplica día a día y pronto demandan prácticamente toda la nutrición disponible para el organismo o para una parte esencial del mismo. En consecuencia, los tejidos normales sufren una muerte nutritiva gradual.
